



Бореальные леса покрывают значительные площади поверхности суши северного полушария, оказывая существенное влияние на региональные и глобальные климатические условия [1]. На территории России сосредоточено более половины всех бореальных лесов планеты. В последнее время исследованию бореальных экосистем северного полушария уделяется повышенное внимание, поскольку для этой зоны прогнозируются наиболее значительные климатические изменения, сопровождаемые ростом температуры, изменением режима осадков и ростом повторяемости аномальных явлений погоды, что, как следствие, может привести к существенным изменениям в структуре и функционировании таежных экосистем. В этой связи особую актуальность получает задача исследования возможных ответных реакций наземной растительности на изменения условий внешней среды (температуру воздуха, газовый состав атмосферы, влагообеспеченность почвы и др.) [2, 3]. Метаболический отклик на воздействие факторов внешней среды у отдельных видов растений зависит как от интенсивности и направленности этих воздействий, так и от физиологических характеристик самих растений [4, 5]. Поэтому межвидовая оценка адаптивного потенциала растений представляет особый интерес при прогнозировании их возможного отклика на изменения природной среды и климата. Следует отметить, что отклик растений на изменения внешних условий проявляется в первую очередь в изменении интенсивности протекания их основных физиологических процессов, прежде всего углеродного и водного обмена. Для прогноза возможной реакции различных растительных сообществ на внешние воздействия необходимо оценить масштабы природной изменчивости, а также чувствительность этих процессов к различным внешним факторам. Полученные показатели могут быть использованы в качестве параметров в математических моделях продукционного процесса и водного цикла наземных экосистем [6, 7].
Цель нашей работы – определение количественных значений параметров углеродного и водного обмена сосны обыкновенной (Pinus sylvestris L.) и березы повислой (Betula pendula Roth), как наиболее широко представленных видов древесных растений в условиях среднетаежной зоны Северо-Запада России, и оценка возможных изменений потоков CO2 и H2O в сосновых лесных экосистемах Карелии при изменении климатических условий в будущем по результатам модельных расчетов.
Материалы и методы исследования
Объекты и методика измерений СО2/Н2О-обмена растений. Полевые исследования СО2/Н2О-обмена проводились в сосняке черничном свежем европейской части средней тайги (Южная Карелия, 62°13′N, 34°10′Е). Состав древостоя 9С1Б, где С – сосна, Б – береза, II–III класс бонитета – возник после беглого пожара в середине прошлого века. Почва – песчаный иллювиально-гумусово-железистый подзол. Основная масса корней сосредоточена в почвенной толще от 2 до 20 см. Грунтовые воды залегают на глубине 1,5–1,7 м [8].
Объектами исследования в зависимости от поставленных задач были подрост и деревья сосны обыкновенной (Pinus sylvestris L.) и березы повислой (Betula pendula Roth) без внешних признаков угнетения. Исследования параметров углеродного и водного обмена деревьев сосны и березы проводились в течение вегетационных периодов 2007–2017 гг. Для измерения показателей СО2/H2O-обмена растений в исследованиях использовалась фотосинтетическая система Li-6400ХТ (Li-Cor Inc., США) и камера давления Plant Moisture Vessel SKPM 1400 (Skye Instruments Ltd., Великобритания). Программа измерений параметров СО2-газообмена в полевых условиях включала получение углекислотных и световых кривых фотосинтеза при разных температурах воздуха и температурных зависимостей темнового дыхания [5]. Для исследования вегетационной динамики суточного ритма водного потенциала охвоенных (облиственных) побегов сосны и березы отбор образцов проводили с интервалом в 1 час в течение суток с июня по сентябрь в дни с разными погодными условиями. В каждый срок наблюдения проводили для 6 деревьев; с каждого дерева отбирали по 3 побега со средней части кроны. Для регистрации интенсивности света, температуры и относительной влажности воздуха, дефицита упругости водяного пара использовали стандартные метеорологические приборы, систему Li-6400ХТ (Li-Cor Inc., США) и автономную систему непрерывной регистрации температуры и относительной влажности ТРВ-2 (Инженерные технологии, Россия). Исследования выполнены на научном оборудовании Центра коллективного пользования Федерального исследовательского центра «Карельский научный центр Российской академии наук».
Для обработки экспериментальных данных использовали корреляционный, регрессионный и дисперсионный анализы. Проверку гипотез и оценку существенных различий между средними величинами осуществляли с помощью критерия Стьюдента и критерия Фишера при 5 %-ном уровне значимости. Статистическую обработку и анализ данных проводили с использованием программ Microsoft Excel и Statistica v.10.
Описание модели и сценарии модельных экспериментов. Для проведения модельных экспериментов по оценке возможных изменений потоков CO2 и H2O в сосновых лесных экосистемах при изменении климатических условий была использована модель MixFor-SVAT [7]. Одномерная процесс-ориентированная математическая модель MixFor-SVAT была разработана для расчета потоков радиации, тепла, Н2О и СО2 как в монодоминантных, так и в смешанных древостоях. В качестве основной концепции модели используется сопряженное описание физико-биологических процессов, протекающих в лесной экосистеме от уровня отдельного листа до уровня отдельного дерева, древостоя и всей экосистемы в целом. Модель состоит из тесно взаимосвязанных блоков, описывающих: перенос солнечной радиации; атмосферной турбулентности; транспирацию; испарение задержанных растительностью атмосферных осадков; испарение с поверхности почвы; фотосинтез, гетеротрофное и автотрофное дыхание растений и почвы; перенос тепла и влаги в почве. Основным преимуществом модели является ее способность не только определить интегральные потоки Н2О и СО2 на уровне всей экосистемы, но также и адекватно оценить вклад почвы, лесной подстилки, а также деревьев разных пород в суммарные потоки парниковых газов с учетом их индивидуальной реакции на изменения условий внешней среды, фенологии, а также различий в структуре и биофизических свойствах [9].
Верификация модели была проведена в ходе многочисленных экспериментов, с использованием экспериментальных данных в различных типах лесных экосистем, и в том числе в сосняках автоморфного ряда в условиях Южной Карелии, и показала ее способность адекватно описывать атмосферные потоки и их изменчивость в лесных экосистемах в широком спектре метеорологических условий.
Для прогноза будущих климатических условий были использованы расчеты по глобальной климатической модели ECHAM5 для сценария А1В [10]. Модельные эксперименты были проведены для двух сценариев возможных изменений растительности. Первый сценарий предусматривал сохранение современной структуры древостоя в будущем. Второй сценарий предполагал изменение надземной и подземной биомассы деревьев (величины листового индекса LAI, диаметра стволов и плотности тонких корней деревьев), рассчитываемой в зависимости от изменения величины первичной нетто продуктивности (NPP) древостоя при будущих климатических условиях. Оба сценария предполагали наличие достаточного количества доступного почвенного азота как при современных условиях, так и в будущем.
Результаты исследования и их обсуждение
Параметры водного обмена. Проведенное исследование влияния интенсивности ФАР, температуры и дефицита упругости водяного пара на вегетационную динамику суточного ритма водного потенциала побегов хвойного и лиственного видов древесных растений в условиях сосняка черничного свежего позволило построить множественную регрессионную модель взаимосвязи водного потенциала сосны и березы от ключевых параметров внешней среды:
ψ = а0 + а1•Т + а2•VPD + а3•PAR,
где ψ– водный потенциал (МПа); Т – температура воздуха ( °С); VPD – дефицит упругости водяного пара (кПа); PAR – интенсивность ФАР (мкмоль/м2·с); а0–а3 – коэффициенты модели (табл. 1).
Рассчитанные стандартизированные коэффициенты регрессионной модели были сопоставлены друг с другом для последующего ранжирования соответствующих им факторов по значимости воздействия на исследуемую переменную. Анализ модельных данных показал, что вклады выделенных внешних факторов в изменчивость величины ψ для сосны и березы хорошо сопоставимы, и межвидовые различия незначительны. Так, например, установлено сходство доминирования ведущих факторов для обоих видов, в частности в июле – интенсивности ФАР, августе – дефицита влажности воздуха, сентябре – температуры воздуха. Однако в июне нами были выявлены значимые межвидовые отличия (p < 0,05) вклада средообразующих факторов в формирование величины ψ, которые зависели у сосны и березы от VPD и PAR соответственно. Этот факт, вероятно, обусловлен биологическими особенностями роста побегов и развития хвои сосны и листьев березы, в частности формирования ассимиляционной и транспирирующей поверхности в период интенсивного роста растений.
Таблица 1
Коэффициенты регрессионной модели взаимосвязи водного потенциала побегов сосны обыкновенной (над чертой) и березы повислой (под чертой) с параметрами внешней среды
|
Месяц |
a0 |
a1 |
a2 |
a3 |
R2 |
|
Июнь |
–1,631*** –0,664*** |
0,017*** ns |
0,011*** 0,004*** |
–0,375*** –0,330*** |
0,89* 0,87* |
|
Июль |
–0,604*** –0,115*** |
–0,012*** –0,009*** |
0,004*** ns |
–0,349*** –0,384*** |
0,85* 0,86* |
|
Август |
–1,951*** –2,664*** |
0,017*** ns |
0,014*** 0,025*** |
–0,305*** –0,287*** |
0,82* 0,81* |
|
Сентябрь |
–1,130*** –0,880*** |
0,017*** 0,017*** |
0,006*** 0,005*** |
–0,231*** –0,309*** |
0,79* 0,74* |
Примечание: R2 – коэффициент детерминации; * p < 0,05, ** p < 0,01, *** p < 0,001, ns – незначимо (p > 0,05).
Следует также отметить некоторую инертность отклика водного потенциала исследуемых растений на резкие изменения абиотических факторов в течение суток, что обусловлено механизмом эндогенной регуляции водного обмена. Так, при определенной величине ψ начинается устьичное ограничение процесса транспирации, что снижает интенсивность дегидратации водопроводящих путей. Степень и интенсивность такой гидравлической регуляции варьирует среди видов и зависит от возраста, структуры и функционального типа растения [4, 3]. Критический порог потери влаги также связывают с величиной ψ, индуцирующего потерю гидравлической проводимости ксилемы ствола хвойных и лиственных древесных растений на 50 и 80 % соответственно [11]. Согласно нашим данным [8] у сосны обыкновенной ограничение транспирационных расходов влаги посредством закрытия устьиц начинается при значениях ψ –1,0…–1,1 МПа. Снижение оводненности ствола у березы повислой происходит при более высоких значениях ψ облиственного побега [12], поскольку диапазон величин ψ, которому соответствует максимальная скорость водного потока у березы, сдвинут в сторону более высоких значений (–0,4…–0,6 МПа) по сравнению с сосной (–0,6…–0,95 МПа). Также анализ наших многолетних данных суточной динамики водного потенциала охвоенных (облиственных) побегов хвойного и лиственного видов показал постоянство диапазонов их предрассветных (ψпоб.max) и дневных (ψпоб min) значений, которые составили для сосны –0,3…–0,7 и –0,65…–1,5 МПа и березы –0,1…–0,45 и –0,5…–1,6 МПа соответственно.
Параметры СО2-газообмена. Для характеристики СО2-газообмена листа мы использовали классический подход Фаркхара [13], предполагающий, что нетто фотосинтез листа зависит от скорости карбоксилирования рибулозобифосфаткарбоксилазы/оксигеназы (РБФК/О) (Vcmax), скорости транспорта электронов при световом насыщении для регенерации акцептора рибулозо-бифосфата (Jmax), темнового дыхания (Rd) и фактора, характеризующего доступность внутренних неорганических фосфатов для цикла Кальвина (TPU).
На основе полученных в ходе измерений углекислотных кривых СО2-газообмена согласно методике, описанной Шаркей с соавторами [14], были рассчитаны значения максимальной скорости карбоксилирования РБФК/О, скорости транспорта электронов для регенерации акцептора РБФ при световом насыщении и скорости утилизации триозофосфатов. Температурные зависимости Vcmax, Jmax и TPU были получены путем статистического анализа множества значений параметров фотосинтеза и дыхания при разных температурах листа. На основании температурных зависимостей были получены значения Vcmax, Jmax и TPU для выбранной референтной температуры 25 °С. Результаты расчета параметров фотосинтеза показали, что хвоя сосны характеризуется более низкими значениями Vcmax, Jmax и TPU по сравнению с листом березы (рисунок). В качестве рабочей гипотезы можно сделать предположение, что более высокие значения скорости фотосинтеза у лиственных пород могут быть связаны с более эффективной работой их фотосинтетического аппарата.
Оценка потоков СО2 и Н2О в лесных экосистемах при будущих изменениях климата. Результаты полевых исследований составляющих СО2/Н2О-обмена хвойных и лиственных видов древесных растений в разных условиях произрастания [8, 12, 15] были использованы для параметризации скорости фотосинтеза, дыхания и устьичной проводимости сосны и березы в модели MixFor-SVAT и проведения расчетов возможных изменений потоков CO2 и H2O в сосновых лесных экосистемах Карелии при изменении климатических условий в будущем (табл. 2).
Результаты модельных экспериментов показали, что прогнозируемый рост температуры и увеличение количества осадков, сопровождаемое возможным изменением видового состава и общей фитомассы растительности может оказать существенное влияние на величины нетто экосистемного обмена СО2 (NEE), валовой первичной продукции (GPP), нетто первичной продукции (NPP) древостоев и экосистемного дыхания (RE) в условиях Северо-Запада России. На фоне роста GPP, RE и NEE прогнозируемое относительное увеличение годового испарения (E), несмотря на прогнозируемый рост температуры и количества осадков, будет довольно незначительным. Это может быть обусловлено прогнозируемым снижением приходящей солнечной радиации, которое, несмотря на увеличение продолжительности вегетационного периода и температуры, может вызвать даже незначительное снижение транспирации (TR) древостоя. Прогнозируемое увеличение влагосодержания почвы и поверхностного стока, за счет роста к концу XX в. количества осадков и незначительного изменения испарения, может также привести к снижению влияния атмосферных засух на лесные экосистемы.
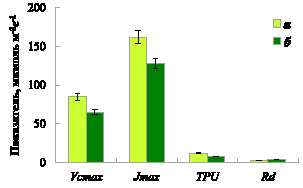
Параметры модели фотосинтеза (Vcmax – скорость карбоксилирования РБФК/О, Jmax – скорость транспорта электронов для регенерации акцептора РБФ при световом насыщении, TPU – скорость утилизации триозофосфатов, Rd – скорость темнового дыхания) березы повислой (а) и сосны обыкновенной (б) при температуре листа 25 °С
Таблица 2
Современная и спрогнозированная на конец XXI в. (ECHAM5, SRES A1B) годовая изменчивость E, TR, NEE, GPP, RE и NPP соснового леса подзоны средней тайги европейской территории России при современных и прогнозируемых будущих климатических условиях на конец XXI в.
|
E |
TR |
NEE |
GPP |
RE |
NPP |
|
|
(мм год-1) |
(г C м-2 год-1) |
|||||
|
Современные условия |
418,5 |
263,7 |
–312,9 |
1441,1 |
1128,2 |
599,1 |
|
Сценарий A1B |
437,6 |
247,3 |
–433,7 |
1895,2 |
1461,5 |
787,8 |
Заключение
Проведенные эксперименты показали высокую чувствительность составляющих СО2/Н2О-обмена сосны обыкновенной (Pinus sylvestris L.) и березы повислой (Betula pendula Roth) к изменению факторов внешней среды, что позволило спрогнозировать возможные будущие изменения составляющих углеродного и водного баланса сосновых и березовых древостоев при разных сюжетных линиях климатических изменений в XXI в. Поддержание экологического равновесия растительного сообщества на фоне изменения СО2 в атмосфере требует строго сбалансированного поступления и потребления элементов минерального питания, воды, света для продукционного процесса. Любой дисбаланс этого комплекса факторов может привести к нарушению устойчивости и повлиять на продуктивность растений. Это обуславливает важность проводимых исследований для построения более точных прогнозов динамики лесных экосистем в будущем и оценки роли лесов в глобальной климатической системе.
Финансовое обеспечение исследований осуществлялось из средств федерального бюджета на выполнение государственного задания КарНЦ РАН (тема № 0220-2017-0002) и при финансовой поддержке РФФИ (грант 17-04-01087-а).
Библиографическая ссылка
Придача В.Б., Ольчев А.В., Сазонова Т.А., Тихова Г.П. ПАРАМЕТРЫ СО2/Н2О-ОБМЕНА ДРЕВЕСНЫХ РАСТЕНИЙ КАК ИНСТРУМЕНТ МОНИТОРИНГА И ОЦЕНКИ СОСТОЯНИЯ ПРИРОДНОЙ СРЕДЫ // Успехи современного естествознания. 2019. № 11. С. 25-30;URL: https://natural-sciences.ru/ru/article/view?id=37235 (дата обращения: 17.05.2026).