


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
DYNAMICS MORFOFUNCTION OF INDEXS OF A FLYING MUSCULATION OF BEES IN AN ONTOGENESIS

Летная активность пчел обеспечивается работой асинхронной летательной мускулатуры, которая представляет собой наиболее совершенный вариант строения мускулатуры беспозвоночных. Функциональной единицей здесь является многоядерное мышечное волокно, образующееся при эмбриональном развитии в результате слияния отдельных клеток. В основе сокращения лежат конформационные изменения внутриклеточных белковых комплексов - актина и миозина. В результате их взаимодействия происходит гидролиз молекул АТФ и химическая энергия превращается в механическую работу, которая реализуется в мышечном сокращении. С точки зрения интенсивности работы (использование максимального количества АТФ в единицу времени) асинхронные летательные мышцы насекомых представляют собой наиболее рациональную механохимическую сократимую систему. Высокая эффективность работы контрактильного аппарата обеспечивается его ультраструктурой.
С целью изучения особенностей структурной организации мышечных волокон в онтогенезе, проводили исследование летательной мышцы медоносной пчелы с использованием световой и электронномикроскопической техники. Для исследования ультраструктуры мышечных тканей, их кусочки объемом 1 мм2, а также отдельные изолированные мышечные волокна фиксировали в 1%-ном растворе ОsО4 на веронал-ацетатном буфере (рН 7,4) по Колфилду и после стандартной проводки через спирты и ацетон заливали в аралдит или эпон-аралдит. Готовили полутонкие и ультратонкие срезы, контрастировали нитратом свинца и просматривали в электронном микроскопе JЕМ-100. Получение мерных показателей органелл (объемные доли, площади поверхностей, их число в единице объема цитоплазмы) проводили по методике Э.Р.Вейбеля (1970) и Р.К.Данилова, В.Н.Сперанского (1988) непосредственно с негативов, увеличенных с помощью увеличителя «Беларусь» при Ку = 3, 5. Активность АТФ-азы определяли по методу Падикула и Герман при рН 9,4. Активность сукцинатдегидрогеназы определяли тетразолиевым методом по Нахласу.
По результатам электронномикроскопических исследований, на стадии куколки площадь мышечного волокна равна 79,0 мкм2. Диаметр миозиновых миофиламентов в этот период составляет 122 А, длина саркомера - 1,91 мкм. Суммарный объем митохондрий в симпластах куколки составил 14% от общего объема клетки. При этом доля митохондрий, расположенных по периферии клетки в 2,13 раза превышала долю интерфибриллярных митохондрий.
У пчел 1-3 суточного возраста площадь мышечных волокон достоверно увеличивается и составляет 92,0 мкм2. Диаметр миозиновых миофиламентов за этот период возрастает до 140 А, длина саркомера - до 2,26 мкм. Соотношение периферических и интерфибриллярных митохондрий в мышечных волокнах на этой стадии развития выравнивается. Суммарное число митохондрий, хотя и превышает этот показатель на стадии куколки, однако остается еще невысоким и составляет 25% (рис.1). Окислительно-восстановительные процессы на кристах митохондрий также не достигают еще уровня летных пчел, о чем свидетельствует низкая активность сукцинатдегидрогеназы - 0,18 усл.ед. На этой стадии зарегистрирован минимальный для имаго показатель гистохимической активности миозиновой и митохондриальной АТФ-аз, ответственных за расщепление макроэргических связей в молекуле АТФ и обеспечение энергией процессов мышечного сокращения - 1,12 и 0,52 усл.ед. соответственно.
Морфометрические и гистохимические показатели летательной мышцы рабочих пчел 4-7 суточного возраста свидетельствуют о значительной физиологической перестройке. У пчел этой возрастной группы площадь мышечного волокна составляет 106 мкм2, диаметр миозиновых миофиламентов 175 А, а длина саркомера 2,55 мкм, что превышает соответствующие показатели на стадии куколки соответственно в 1,34; 1,43 и 1,33 раза.
Число митохондрий в продольной грудной мышце возрастает и составляет уже 36% от объема симпласта. Соотношение периферических и интерфибриллярных митохондрий меняется в сторону преобладания последних (1:2,52). Активность СДГ у пчел этой возрастной группы составляет 2,43 усл.ед., что выше показателя предыдущей группы в 13,5 раза. Гистохимическая активность АТФ-аз миозина и митохондрий регистрируется на уровне 3,3 и 2,33 усл.ед. Достигнутый на этой стадии уровень обменных процессов уже достаточен для обеспечения кратковременного полета. Именно в этом возрасте нами регистрировался вылет меченых рабочих пчел на первые очистительные и ориентировочные облеты (рис.2).
В возрасте 10-12 суток площадь мышечных волокон по сравнению с куколкой увеличивается в 1,39 и составляет 110 мкм2. Диаметр миозиновых нитей на этой стадии - 186 А, длина саркомера - 2,7 мкм. Общее число митохондрий существенно возрастает, они занимают 55% объема мышечного волокна. В этот период наблюдается первый максимальный пик преобладания интерфибриллярных митохондрий, отношение их к периферическим составляет 3,45:1. Активность СДГ повышается в 18,94 раза по сравнению с первой имагинальной группой - до 3,41 усл.ед.
Значительное повышение метаболической активности летательной мускулатуры наблюдается у пчел 11-20 суточного возраста. Хотя общее число митохондрий увеличивается незначительно и составляет 58%, а соотношение интерфибриллярных и периферических несколько снижается, в то же время, активность окислительно-восстановительных процессов на кристах митохондрий резко возрастает. Так, уровень СДГ повышается до 8,19 усл.ед. и превышает уровень 1-3 суточных пчел в 45,5 раза. В этот период пчелы начинают вылетать для сбора нектара и летная активность нами регистририровалась у 80% меченых пчел.
Готовность летательной мускулатуры к длительному полету проявляется положительной динамикой морфометрических показателей структурных единиц мышечного волокна и нарастанием активности ферментов, обеспечивающих энергией процессы сокращения. В возрасте 11-20 суток площадь мышечных волокон по сравнению с куколкой увеличивается в 1,46 раза и составляет 115 мкм2. Диаметр миозиновых нитей возрастает до 222, длина саркомера - до 2,75. Активность АТФ-азы миозина у пчел 11-20 суточного возраста достигает 6,52 усл.ед., митохондриальной АТФ-азы - 15,24 усл.ед., то есть превышает показатель первой имагинальной стадии соответственно в 5,82 и 29,3 раза.
У меченых пчел после 20-го дня жизни при активном сборе нектара морфофункциональные показатели летательной мышцы стабилизируются. Некоторое снижение к 21-30 суточному возрасту наблюдается лишь для площади мышечного волокна и диаметра миозиновых миофиламентов. Эти показатели составляют соответственно 114 мкм2 и 221 А. В динамике длины саркомера по сравнению с предыдущей стадией наблюдается незначительное увеличение - до 2,8 мкм. Заметно возрастает активность миозиновой АТФ-азы - до 9,35 усл.ед., а митохондриальной остается практически на прежнем уровне - 15,31 усл.ед. Данный период жизни пчел характеризуется активной летной деятельностью и требует максимальных затрат энергии. На этой стадии регистрируется второй пик максимального преобладания интерфибриллярных митохондрий над периферическими. Их соотношение в симпласте составляет 3,78:1. Активность СДГ повышается по сравнению с первой имагинальной стадией в 48 раз и составляет 8,65 усл.ед.
Следовательно, в процессе миогистогенеза происходят качественные и количественные перестройки контрактильного и энергетического аппаратов, связанные с терминальной дифференцировкой мышечных волокон. При этом существенно повышается роль интерфибриллярных митохондрий, обеспечивающих высокую ферментативную активность в дефинитивных мышечных волокнах и адаптацию мышечной ткани к возрастанию нагрузки.
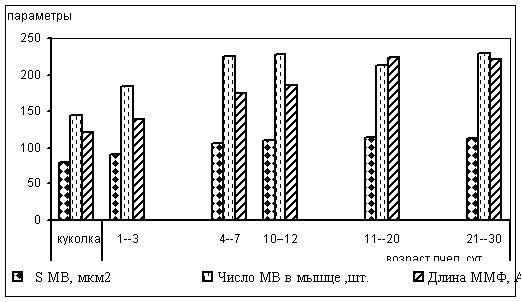
Рисунок 1. Основные параметры мышечного волокна летательной мускулатуры пчел в онтогенезе (МВ - мышечное волокно, ММФ - миозиновые миофиламенты)
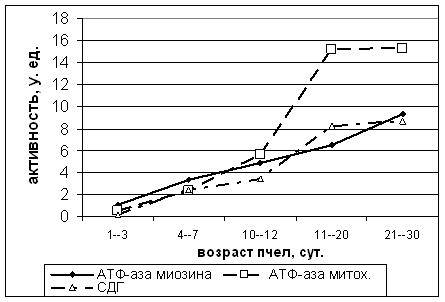
Рисунок 2. Показатели активности АТФ-азы миозина, АТФ-азы митохондрий и СДГ а летательной мышце пчел
СПИСОК ЛИТЕРАТУРЫ
- Альбертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж.. Молекулярная биология клетки. - М. Мир 1970. С.296.
- Басурманова О.К., Бочарова-Месснер О.М. // Молекулярная и клеточная биофизика. М. Наука. 1977. C.210-216
- Бендолл Дж. Мышцы, молекулы и движение. М., Мир, 1970. С.256.
- Смит Д. // Молекулы и клетки. М., Мир, 1970, C.94-107.
- Auber I. // Amer.Zoologist. 1967. № 7. P.451-456 .
- Franzini-Armstrong C. // The structure and function of muscles. 1973. 2. N.Y.- London. P. 532-620.
Библиографическая ссылка
Мишуковская Г.С., Маннапов А.Г. ДИНАМИКА МОРФОФУНКЦИОНАЛЬНЫХ ПОКАЗАТЕЛЕЙ ЛЕТАТЕЛЬНОЙ МУСКУЛАТУРЫ ПЧЕЛ В ОНТОГЕНЕЗЕ // Успехи современного естествознания. 2004. № 8. С. 19-22;URL: https://natural-sciences.ru/en/article/view?id=13217 (дата обращения: 07.07.2026).