



![]()
Также исследован процесс возрастания энтропии в твердом теле, в котором имеется градиент температур [3]. По определению изменение удельной энтропии равно
![]()
Используя для рассмотрения указанного процесса закон сохранения энергии в дифференциальной форме, имеющий вид уравнения неразрывности, было получено следующее выражение для производства энтропии [3]
![]()
где W¯ вектор плотности потока энергии. Выражение
![]()
- источник искомого приращения энтропии в единице объема в единицу времени.
В настоящей работе рассмотрено производство энтропии в адиабатически замкнутой системе, в которой наряду с градиентом температуры существуют внутренние источники тепла с производительностью q (Вт/м3).
Изменение удельной энтропии по-прежнему характеризуется выражением (2), а закон сохранения энергии в дифференциальной форме имеет вид уравнения теплопроводности [4]
![]()
которое можно преобразовать к виду
![]()
где λ = a·c·p - коэффициент теплопроводности.
Учитывая что ![]() сравнивая (3) и (6), получаем
сравнивая (3) и (6), получаем
![]()
Воспользуемся тождеством
![]()
или
![]()
Сравнивая (9) и (7), получаем
![]()
Проинтегрируем (10) по всему объему замкнутой системы
![]()
Согласно условиям решаемой задачи и по теореме Гаусса-Остроградского получим
![]()
Итак, имеем
![]()
Сравнивая выражения (3) и (4), отмечаем, что первый интеграл в правой части выражения (13) определяет приращение энтропии, связанное с наличием градиента температур, а второй интеграл в правой части выражения (13) определяет производство энтропии, связанное с производством тепла внутренними источниками.
В дифференциальной форме производство энтропии имеет следующий вид
![]()
В линейной области выражение для производства энтропии определяется выражением [3]
![]()
где ![]() термодинамические потоки,
термодинамические потоки,
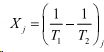 - термодинамические силы.
- термодинамические силы.
С учетом (1), (15) выражение (14) может быть записано в виде:
![]()
где T1 - температура внеутреннего источника тепла,
Т2 - температура среды,
T1-T2 =δT1
C - удельная теплоемкость.
Как следует из (16), при ![]() временное измене-ние производства энтропии запишется в виде:
временное измене-ние производства энтропии запишется в виде:
![]()
Очевидное соотношение, определяющее выделение тепла внутренними источниками ![]() запишется в виде
запишется в виде
![]()
после преобразования получаем
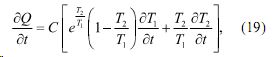
Отток тепла от внутреннего источника можно записат также в виде:
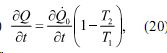
где Q[c точкой] 0 - удельная теплопродукция при ![]()
Исходя из представлений равновесной термодинамики о сохранении баланса тепла, которое выполняется при 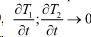 , можно записать
, можно записать
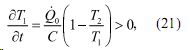
что в свою очередь приводит к получению условия устойчивости термодинамической системы
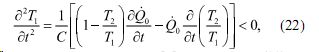
При этом, как показано в (5), выражение (22) преобразуется к виду:
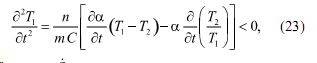
где .jpg) коэффициент теплоотдачи, m - масса ограниченная поверхностью системы, n - поверхность системы.
коэффициент теплоотдачи, m - масса ограниченная поверхностью системы, n - поверхность системы.
Как указано в работе [7] производство энтропии в живом организме в состоянии покоя близком к слабонеравновесному характеризуется выражением
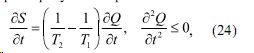
где T1 и T2 - температура живого организма и внешней среды, соответственно,
![]() - величина полной теплопродукции живого организма
- величина полной теплопродукции живого организма
При этом ![]() производство энтропии в живом организме равно продукции энтропии во внешнюю среду.
производство энтропии в живом организме равно продукции энтропии во внешнюю среду.
Одновременно заметим, что при T1 -T 2=δT< T1, что соответствует условиям обитания живых организмов, выражение (24) может быть записано в виде
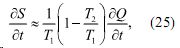
позволяющее заключить, что производство энтропии живых организмов в состоянии покоя достигается путем диссипации тепла при температуре тела живого организма T1.
С другой стороны, величина изменения теплопродукции может быть записана в виде [5,6]
![]()
где α - коэфиициент теплоотдачи живого организма к внешней среде,
П - поверхность тела живого организма.
В работе [7] доказано что величина α(t) есть убывающая функция времени, т. е. ![]() Из выражения (26) следует, что возможно сохранение величины
Из выражения (26) следует, что возможно сохранение величины ![]() . Тогда
. Тогда
что существенно для живого организма при сохранении состояния покоя не равного равновесному, из которого живой организм уже не может удалиться.
Таким образом, из выражений (24-27) следует, что возможно длительное существование в состоянии покоя, при этом ![]() , т. е. достигается состояние температурного гомеостаза и гомеостатирования среды обитания.
, т. е. достигается состояние температурного гомеостаза и гомеостатирования среды обитания.
В филогенезе живых организмов величина П ограничена предельной массой (M) тела живого организма, т. к. известно, что отношение м/п есть величина постоянная, например, для гомойотермных организмов [5,6], что в конечном счете приводит к нарушению функций органов живого организма и его вырождению.
Однако из вышеуказанного следует, что если отношение м/п = const выполняется, или, например, значительному увеличению П соответствует незначительное увеличение M, длительность развития с сохранением практически постоянно удельной теплопродукции может быть значительной.
В живом организме есть орган, для которого возможно выполнение данного условия - это мозг живого организма, значительному изменению поверхности которого соответствует значительно меньшее, чем для других органов тела, увеличение массы.
Этот путь эволюционного развития и использовали живые организмы, но больше всех преуспел человек, который смог в дополнение к температурному гомеостазу создать условия гомеостатирования внешней среды (жилища, одежда, применение огня), что в состоянии покоя привело к формированию мозга, позволяющего вести рассудочную деятельность.
Вышеизложенное составляет сущность термодинамических принципов эволюции живых организмов.
Выводы
Сущность необходимых термодинамических принципов эволюции живых организмов заключается в том, что, как доказано в статье, величина коэффициента теплоотдачи живого организма в филогенезе и онтогенезе является убывающей функцией времени в состоянии покоя, при постоянных температурах тела и внешней среды и для поддержания неизменной полной теплопродукции необходимо увеличение поверхности тела живого организма, что определяет удаление живого организма от состояния покоя.
При невыполнении в отдельном органе живого организма - мозге, известной в физиологии зависимости массы всего живого организма от его поверхности, например для гомойотермных организмов, возникает увеличение массы и поверхности мозга, позволяющее живым организмам, при достижении гомеостатирования внешней среды, эво-люционизировать путем формирования мозга не снижая удельной теплопродукции, что подтверждается эволюцией гомойотермных организмов.
Если формирование мозга отсутствует, то, при достижении гомеостатирования внешней среды, эволюция живых организмов приводит к увеличению поверхности тела живых организмов и, соответственно, к пропорциональному увеличению их массы, при этом происходит снижение удельной теплопродукции, что подтверждается гибелью гиперфауны.
Выживание наиболее приспособленных живых организмов при воспроизводимых условиях внешней среды не связано с достижением ими состояния покоя и с изменением удельной теплопродукции в филогенезе, что подтверждается конкуренцией биоценозов и составляет сущность идиоадаптации.
Полученные результаты позволяют выполнить термодинамическое обоснование пунктуализма эволюционного процесса, известного как ароморфоз Северцева, т. е. показать, что пунктуализм изменений связан с периодическим достижением гомеостатирования среды обитания живых организмов, при этом прогрессивные изменения в живом организме, возникают как результат противодействия состоянию покоя при гомеостатировании среды обитания.
Необходимо отметить, что человек достиг состояния температурного гомеостаза и гомеостатирования среды обитания в состоянии покоя, что привело к прогрессивному увеличению его размеров.
БИБЛИОГРАФИЧЕСКИЙ СПИСОК
- Левич, В. Г. Введение в статистическую физику / В. Г. Левич. - М.: Гостехиздат, 1954. - С. 141.
- Волькенштейн, М. В. Биофизика / М. В. Волькен-штейн. - М.: Наука, 1988. - С. 16-19, 307-308, 566-573.
- Ансельм, А. И. Основы статистической физики и термодинамики / А. И. Ансельм. - М.: Наука, 1973. - С. 355357.
- Исаченко, В. П. Теплопередача / В. П. Исаченко, В. А Осипова, А. С. Сукомел. - М.: Энергия, 1969. - С. 15-18,27-28.
- Шмидт-Ниельсон, К. Размеры животных: почему они так важны / К. Шмидт-Ниельсон. - М.: Мир, 1987. - 259 с.
- Проссер, Л. Сравнительная физиология животных: Пер. с англ. / Л. Проссер, Ф. Браун. - М.: Мир, 1967. - С. 11-123.
- Кузнецов, В. Г. Производство энтропии в адиабатической замкнутой системе с градиентом температуры и производством тепла и применение этих условий к термодинамической модели живых организмов / В. Г. Кузнецов // Биофизика. - 2003. - Т. 48. - Вып. 3. - С. 572-573.