


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
THE ROLE OF MONOAMINERGIC CNS SYSTEMS IN DISPLAYING OF ADAPTIVE CHANGES IN ORGANISM

Однако, как показывает практика, после прекращения сеансов адаптации, сформированный адаптивный защитный след существует относительно недолго, он постепенно «стирается» и после этого наступает дезадаптация. Поэтому для поддержания системного структурнофункционального следа адаптации, обеспечивающего надежную профилактику или целенаправленную коррекцию тех или иных функциональных нарушений у человека необходимо проводить регулярные сеансы адаптации людей к условиям гипоксии. Такой подход становится достаточно трудоемким и поэтому столь эффективный метод коррекции функциональных возможностей организма не нашел должного распространения в клинике.
Согласно ряду исследований [3, 5, 6] в адаптации организма к тем или иным воздействиям существенную роль играют нейротрансмиттерные системы, воздействуя на которые можно пролонгировать, сформированный адаптивный след. Поэтому в настоящей работе исследованы медиаторные механизмы, обеспечивающие адаптивные перестройки в организме. Цель исследования заключалась в изучении состояния моноаминергических (МА-) систем различных отделов мозга после адаптации организма к периодической гипоксии.
Исследования выполнены на стандартных белых крысах-самцах линии Вистар массой 160 -180 г. Концентрацию моноаминов (МА) в структурах ЦНС определяли с помощью метода высокоэффективной жидкостной хроматографии с электрохимической детекцией [1]. Адаптацию животных к гипоксии проводили в барокамере на «высоте» 5000 м в течение 30 суток по 6 часов ежедневно.
Результаты исследований показали, что после месячной адаптации крыс к периодической гипоксии в условиях барокамеры, существенно меняется характер межмедиаторных взаимоотношений в МА-ергических системах различных отделов мозга (рис.).
Установлено, что через сутки после последнего сеанса адаптации в коре головного мозга существенно увеличивается (на 25 %) уровень норадреналина (НА), тогда как концентрация дофамина (ДА) снижается на 30 %. Полученные экспериментальные данные показали, что снижение уровня ДА могло быть обусловлено активацией процессов, обеспечивающих его превращение в НА, при этом концентрация диоксифенилуксусной кислоты (ДОФУК) оказалась статистически достоверно увеличена на 15 % по сравнению с контролем.
Исследования концентраций серотонина (СТ) установили, что адаптация организма к гипоксии сопровождается накоплением СТ в коре головного мозга, превышающем контрольный уровень на 22 %. Определение одного из метаболитов СТ, а именно 5-оксииндолилуксусной кислоты (5-ОИУК), показало, что увеличение концентрации СТ связано со снижением его деградации. Таким образом, полученные результаты свидетельствуют о том, что после адаптации животных к гипоксии в коре головного мозга обнаруживается активация НА- и СТ-ергической систем.
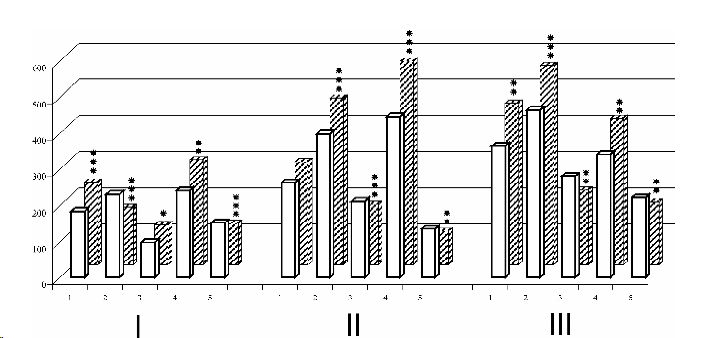
Рисунок 1. Концентрация моноаминов в различных отделах головного мозга животных после адаптации их к гипоксии (в нг/г сырой ткани). 1 - НА; 2 - ДА; 3 -ДОФУК; 4 - СТ; 5 - 5-ОИУК; I - кора; II- средний мозг; III - продолговатый мозг; □ - контроль; ■ - опыт; * - p<0,05; ** - p<0,01; *** -p<0,001.
Для сопоставления уровня медиаторов в коре, где находятся в основном терминали МАергических систем, с отделами мозга, содержащими МА-синтезирующие нейроны, был изучен метаболизм МА в среднем и продолговатом мозге.
Оказалось, что после адаптации концентрация НА в среднем мозге не изменялась, тогда как в продолговатом мозге уровень НА был выше контрольной величины на 24 %. Содержание же ДА в среднем мозге увеличивалось по отношению к контролю на 16 %, что, по-видимому, связано со снижением уровня активности моноаминоксидаз. В пользу этого свидетельствует уменьшение на 20 % одного из метаболитов ДА, а именно ДОФУК.
Как свидетельствуют результаты исследований, адаптация организма к гипоксии приводит к изменению состояния СТ-ергической системы среднего мозга. Так, через сутки после прекращения сеансов адаптации здесь выявлено увеличение концентрации СТ на 27 %. Такие изменения в значительной мере могут быть обусловлены снижением активности ферментативных систем, обеспечивающих превращение СТ в 5ОИУК. В пользу этого свидетельствует снижение концентрации 5-ОИУК в среднем мозге на 30 % после прекращения 30-суточных сеансов адаптации животных в барокамере.
Сопоставление отдельных звеньев метаболизма ДА и НА в среднем и продолговатом мозге обнаружило однотипный характер сдвигов исследованных показателей. Так, у животных, адаптированных к условиям гипоксии, выявлена активация ДА-ергической системы в продолговатом мозге. Концентрация ДА здесь превышала контрольный уровень на 20 %. Однако возможности ДА-синтезирующих структур этого отдела мозга, по-видимому, снижены и накопление запасов ДА в значительной мере связано со снижением активности ферментативных систем, обеспечивающих превращение ДА в ДОФУК и, как следствие, снижение концентрации ДОФУК в среднем мозге. Аналогичный характер сдвигов обнаружен и при исследовании СТ-ергической системы продолговатого мозга. У адаптированных к гипоксии животных содержание СТ увеличивалось на 20 %, при этом содержание 5ОИУК, оказалось статистически достоверно ниже контрольного уровня на 21 %.
Таким образом, проанализировав характер сдвигов в ДА-ергической и СТ-ергической системах среднего и продолговатого мозга у адаптированных к гипоксии животных, можно заключить, что сеансы адаптации активируют эти системы в стволовом отделе мозга. Однако судя по уровню ДОФУК и 5-ОИУК, синтетические возможности ДАи СТ-синтезирующих структур, по-видимому, снижены. Здесь возникает некоторое противоречие между повышенными функциональными потребностями стволовых структур мозга адаптированных животных в дофамине и серотонине и сниженными синтетическими возможностями этих структур мозга. Тогда метаболизм этих МА перестраивается так, что активность процессов деградации ДА и СТ снижается, чтобы сохранить повышенный уровень данных медиаторов, обеспечивающих высокую функциональную активность ДА-и СТ-ергических систем у адаптированных животных.
Исследования показали, что формирование адаптивного следа сопряжено с активацией МАергических систем в исследованных отделах мозга, где после завершения сеансов адаптации выявлено в основном накопление катехоламинов и серотонина. Однако в большинстве случаев оно сопровождалось снижением концентрации продуктов деградации этих МА. Все это свидетельствует о том, что синтетические возможности МА-синтезирующих структур мозга могут быть фактором, лимитирующим адаптивные возможности организма.
СПИСОК ЛИТЕРАТУРЫ
- Бауэр Г., Энгельгард Х., Хеншен А. и др. Высокоэффективная жидкостная хроматография в биохимии. М.: Мир. 1988. 688 с.
- Караш Ю.М., Стрелков Р.Б., Чижов А.Я. Нормобарическая гипоксия в лечении, профилактике и реабилитации. М.: Медицина. 1988. 357с.
- Мамалыга Л.М., Мамалыга М.Л. Роль биогенных аминов в проявлении структурнометаболических сдвигов в ЦНС при стрессе, адаптации и функциональных нарушениях. М. 2004. Изд-во «Прометей». 363 с.
- Меерсон Ф.З., Твердохлеб В.П., Боев В.М., Фролов Б.А. Адаптация к периодической гипоксии в терапии и профилактике. М.: Наука. 1989. 70 с.
- Хайдарлиц С.Х. Нейромедиаторные механизмы адаптации. Кишинев: «Штиинца». 1990.179 с.
- Kvetnansky R., Tilders F. //Endocrinology.1987. V. 45. № 4. P. 318.
Библиографическая ссылка
Мамалыга М.Л., Гуркин А.А. РОЛЬ МОНОАМИНЕРГИЧЕСКИХ СИСТЕМ ЦНС В ФОРМИРОВАНИИ АДАПТИВНЫХ ПЕРЕСТРОЕК В ОРГАНИЗМЕ // Успехи современного естествознания. 2006. № 2. С. 17-19;URL: https://natural-sciences.ru/en/article/view?id=14185 (дата обращения: 16.07.2026).