


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
Postcatelectrotonic potentials and excitability changes following them in nerve fibres

Подробное изучение электротонических потенциалов - физического электротона - осуществлено в 1947 году Р. Лоренте де Но [13], который всесторонне изучив физический кати анэлектротон, выделил в их развитии два компонента: быстрый, длительностью менее 1 мс, и медленный, следующий за быстрым и продолжающийся до выключения поляризующего тока.
В нашей предыдущей работе [5] дополнительно к быстрому и медленному компонентам электротона, развивающимся во время поляризации [13], обнаружен ещё один компонент продолжительный след деполяризации, следующий после медленного электротона постэлектротоническая деполяризация. Целью настоящей работы явилось исследование амплитудно-временных характеристик обнаруженной нами постэлектротонической деполяризации и сопровождающих её изменений возбудимости.
Опыты проводили на одиночных нервных волокнах с «прикрытым» [4] перехватом Ранвье и целых нервах озёрной лягушки. После выдерживания изолированного нерва в растворе Рингера в течение 40 - 90 минут производили выделение из него одиночного нервного волокна.
Препаровку волокна в области перехвата Ранвье производили вместе с прилегающими к нему участками соседних волокон с целью предохранения перехвата от возможного травмирования и растягивания. Исследуемый перехват (N1) помещали в среднюю канавку камеры (рис. 1, А), заполняемую раствором Рингера или растворами изучаемых веществ. Два других перехвата (N1 и N3) с участками нервного ствола располагали на отшлифованных и закруглённых предметных стёклах (рис. 1, А). Активность перехватов N1 и N3 подавляли 0.2% раствором новокаина. Раздражение исследуемого перехвата N2 и отведение от него потенциалов производили с помощью трёх последовательно расположенных неполяризующихся каломельных электродов. Дистальный и средний электроды использовали для нанесения толчков постоянного тока, средний и дистальный - для регистрации ответной реакции. Часть исследований выполнена на математической модели Франкенхёйзера-Хаксли [11].
В опытах на нервном стволе использовали ту же схему стимуляции и регистрации ответа нерва, что и на изолированных нервных волокнах. Отводимые потенциалы подавали на усилитель постоянного тока УУ-2М, затем на осциллограф физиологической установки ЭПМ НИИЭМ АМН СССР и на аналого-цифровой преобразователь компьютера. Нейрограммы обрабатывали с помощью математической программы Mathcad 2000.
Отправным моментом исследования постэлектротонической деполяризации послужили опыты на одиночных перехватах Ранвье изолированных нервных волокон в условиях блокирования натриевых каналов новокаином и тетродотоксином [4]. Было обнаружено, что после приложения к такому нервному волокну деполяризующего электротонического стимула примерно такой же амплитуды и длительности, как и потенциал действия, возникает продолжительный след деполяризации (Рис. 1, В). В процессе ритмической стимуляции нервного волокна данный след суммируется с образованием деполяризационного плато (Рис. 1, Г).
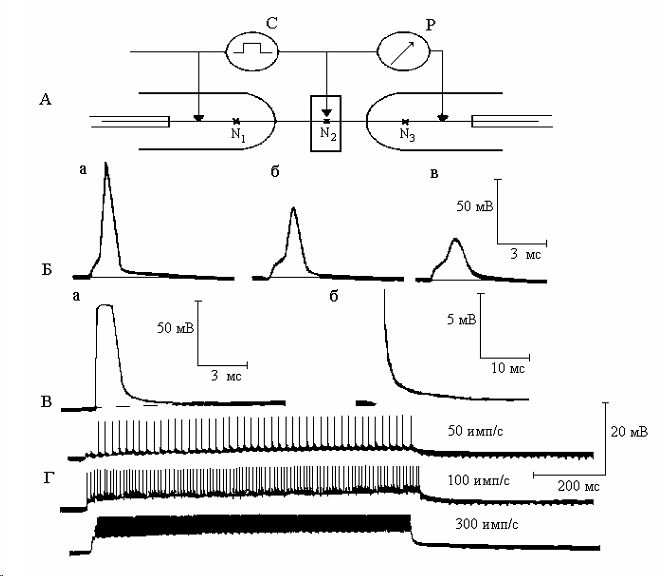
Рис 1.
В опытах на целом нерве для исключения влияния на постэлектротонические потенциалы потенциалов действия, наносили подпорговые катодические стимулы. Оказалось, что после выключения поляризующего тока и прекращения катэлектротонических потенциалов регистрируется аналогичный наблюдаемому на одиночных нервных волокнах след деполяризации. В ответ на приложение к нерву одиночных подпороговых деполяризующих стимулов различной длительности (1,5 и 10 мс) возникал катэлектротонический потенциал (Рис. 2, А) амплитудой 2 3 мВ. После выключения поляризующего стимула происходило постепенное восстановление мембранного потенциала к исходному уровню - постэлектротоническая деполяризация. После приложения катода продолжительностью 1 мс постэлектротоническая деполяризация составила 0.093 ± 0.004 мВ. Снижение постэлектротонической деполяризации происходило по экспоненте в течение 7.123± 0.576 мс. После поляризации нерва катодным током длительностью 5 мс амплитуда и длительность постэлектротонической деполяризации увеличились и составили 0.189 ± 0.005 мВ и 23.212 ± 1.186 мс, а после катэлектротона длительностью 10 мс 0.220 ± 0.011 мВ и 68.721 ± 3.389 мс соответственно.
Для определения изменений возбудимости во время постэлектротонической деполяризации использовали методику нанесения парных стимулов: первого - поляризующего и второго тестирующего, интервал между которыми произвольно изменяли от 1 до 200 мс. Во время постэлектротонической деполяризации обнаружена фаза повышенной возбудимости - потенциал действия в ответ на субмаксимальный тестирующий стимул возрастает по своей амплитуде. Фаза повышенной возбудимости по продолжительности соответствует длительности постэлектротонической деполяризации.
Если на нерв нанести не один, а серию катодических стимулов происходит суммация постэлектротонической деполяризации с образованием небольшого деполяризационного плато (Рис. 2, Б). Величина и характер суммации постэлектротонической деполяризации в процессе ритмической стимуляции зависят от её величины после каждого одиночного стимула. Суммация постэлектротонической деполяризации в наших экспериментах в процессе нанесения серии катодических толчков длительностью 10 мс была более выражена, чем при толчках длительностью 1 мс.
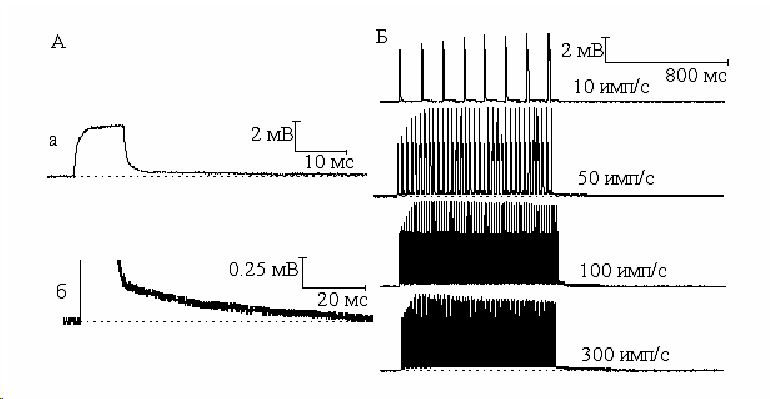
Рис 2.
Таким образом, полученные экспериментальные данные свидетельствуют о том, что выраженность постэлектротонических изменений поляризации и возбудимости зависит от длительности деполяризации мембраны приложением катодического стимула. Постэлектротоническая деполяризация суммируется при ритмическом приложении катодических толчков, причём чем длительнее прикладываемые к нерву катодические толчки, тем выраженнее постэлектротоническая деполяризация.
Возникает вопрос, какие же изменения происходят в нервных волокнах при пропускании через них прямоугольных катодических стимулов. В экспериментах с фиксацией потенциала [8, 9, 12] показано, что в месте приложения прямоугольных деполяризующих стимулов надпороговой силы возникают кратковременно длящийся (около 1 мс) входящий натриевый ток и задержанный выходящий калиевый ток, продолжающийся в течение всей поляризации. Нанесение подпороговых деполяризующих стимулов силой менее 0.5 порога вызывает возникновение только выходящего калиевого тока (рис. 3), а при силах тока, превышающих 0.5 реобазы, возникают локальные токи, состоящие из натриевого и калиевого компонентов. Воспроизведение катэлектротонических потенциалов и сопровождающих их ионных токов осуществлено на математической модели нервного импульса амфибий [11].
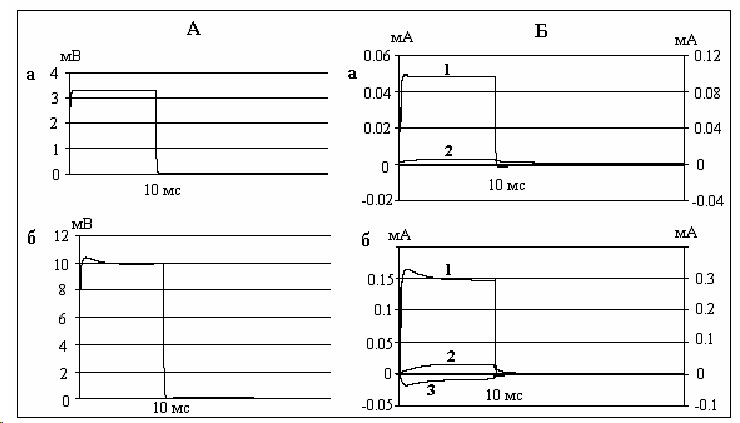
Рис 3.
В проведённых нами опытах с околопороговой катодической поляризацией нерва возникают как натриевый, так и калиевый токи, величина которых в несколько раз меньше по сравнению с аналогичными токами, обусловливающими возникновение потенциала действия. Натриевый ток восходящей фазы локального ответа значительно ослабевает к концу 1 мс. Калиевый же ток продолжается в течение всей деполяризации. Исходя из того, что натриевый ток восходящей фазы локального ответа резко ослабевает к концу 1 мс, а постэлектротоническая деполяризация регистрируется и при длительностях раздражения 5 и 10 мс, реальный вклад ионов натрия в генерацию постэлектротонической деполяризации мало вероятен.
Рост постэлектротонической деполяризации при увеличении амплитуды и длительности катэлектротона, а также наличие продолжительного следа деполяризации у нервных волокон с прикрытым перехватом Ранвье при заблокированных натриевых каналах [4] позволяет рассматривать в качестве наиболее вероятного фактора, определяющего возникновение постэлектротонической деполяризации, выход ионов калия.
Выводы
- После деполяризации возбудимой мембраны одиночного перехвата Ранвье изолированных нервных волокон и целого нерва постоянным током подпороговой силы длительностью 1, 5, 10 мс регистрируется постэлектротоническая деполяризация, представляющая собой медленное восстановление поляризации к исходному уровню. Постэлектротоническая деполяризация одиночных перехватов Ранвье и изолированного нерва обнаруживается не только в исходном состоянии, но и при полном блокировании натриевых каналов.
- Амплитуда и длительность постэлектротонической деполяризации целого нерва при подпороговой деполяризации увеличиваются пропорционально длительности приложенной деполяризации: после пропускания катодического тока продолжительностью 1 мс они составили 0.093 0.004 мВ, а длительность 7.123 0.576 мс, после деполяризации длительностью 5 мс -0.189 0.005 мВ и 23.212 1.186 мс, а после деполяризации длительностью 10 мс 0.220 0.011 мВ и 68.721 3.389 мс соответственно.
- В процессе пропускания через нерв серии катэлектротонических потенциалов происходит суммация постэлектротонической деполяризации, выраженность которой пропорциональна величине и продолжительности поляризации нервных волокон постоянным током.
- На основании наличия постэлектротонической деполяризации у одиночных перехватов Ранвье и изолированного нерва как в исходном состоянии, так и при полном блокировании натриевых каналов, в качестве наиболее вероятного фактора, определяющего возникновение постэлектротонической деполяризации, рассматривается выход ионов калия.
Список литературы
- Вериго Б.Ф. // Труды СПб. Общества естествоиспытателей. - 1883. - Т. 14. - В. 1. - С 15.
- Вериго Б.Ф. К вопросу о действии на нерв гальва3нического тока прерывистого и непрерывного.С.-П. - 1888. (цитировано по: Ходоров Б.И. Общая физиология возбудимых мембран. - М.: Наука, 1975. - 406 С.).
- Воронцов Д.С. // Физиологический журнал СССР им. И.М. Сеченова 1962. - V. 48. № 5. C. 510.
- Каталымов Л.Л. // Доклады АН. 1995. V. 341. № 6. С. 839.
- Каталымов Л.Л., Евстигнеев Д.А. // Успехи соврем. естествознания. 2002. №5. С.45.
- Ходоров Б.И. // Успехи соврем. биол. -1950. Т. 29. - В. 3. - С. 329.
- Ходоров Б.И. // Успехи соврем. биол. -1962. - Т. 54. - В. 3. - С. 333.
- Dodge F.A., Frankenhaeuser B. // J. Physiol.- 1958. - V. 143. - P. 76.
- Dodge F.A., Frankenhaeuser B. // J. Physiol.- 1959. - V. 148. - P. 188.
- Du Bois-Reymond E. Untersuchungen űber thierische Elektrizitat. Berlin. 1848. (цитировано по: Ходоров Б.И., 1975).
- Frankenhaeuser B., Huxley A.F. // J. Physiol. - 1964. - V. 171. - P. 302.
- Hodgkin A.L., Huxley A.F. // J. Physiol. -1952. - V. 117. - P. 500.
- Lorente de No R. A studi of nerve physiology. Stud. Rockfeller Inst. med. Res. 1947. V. 131132. Parts 1-2. 620 С.
- Pflűger E. Untersuchungen űber die Physiologie des Elektrotonus. Berlin. 1859. (цитировано по: Ходоров Б.И., 1975).
Библиографическая ссылка
Евстигнеев Д.А. ПОСТКАТЭЛЕКТРОТОНИЧЕСКИЕ ПОТЕНЦИАЛЫ И СОПРОВОЖДАЮЩИЕ ИХ ИЗМЕНЕНИЯ ВОЗБУДИМОСТИ НЕРВНЫХ ВОЛОКОН // Успехи современного естествознания. 2003. № 6. С. 13-17;URL: https://natural-sciences.ru/en/article/view?id=14434 (дата обращения: 27.06.2026).