


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
CENOPOPULATIONS PYROLA INCARNATA DISTURBED FOREST LENA-AMGA (Central Yakutia)

Pyrola incarnata – циркумполярный бореальный вид, распространен почти во всех областях северного полушария с умеренным и умеренно-холодным климатом. Обладает широкой амплитудой толерантности к абиотическим факторам, таким как увлажнение, освещенность и химический состав почвы.
Центральная Якутия характеризуется равнинным и увалисто-равнинным рельефом с довольно засушливым климатом и холодной продолжительной зимой. Лено-Амгинское междуречье занимает восточную часть Центрально-Якутской равнины, общая площадь лесного фонда 9,1 млн. га, лесистость территории равна 72 %. Здесь господствует светлохвойная тайга из Larix cajanderi (90 %) с незначительным участием Pinus sylvestris (9,5 %) [2].
Pyrola incarnata встречается по всей лесной территории Лено-Амгинского междуречья, где она выступает как один из доминантов в травяно-кустарничковом покрове. В ксерофильных сообществах сосновых и травянистых лиственничных лесов ее участие в травяно-кустарничковом ярусе незначительна, где встречается редкими фрагментами.
Материал и методы исследования
Полевой сбор материала был произведен в июле-августе 2010 и 2011 годов. Геоботанические и популяционные исследования проводили методом закладки трансект и пробных площадей. Закладывались площадки с площадью 1 м2, в среднем по 3 площадки на каждом участке, количество площадок зависело от плотности парциальных образований. В работе использованы общепринятые популяционно-онтогенетические, и геоботанические методы [1, 3].
Исследования особей и ЦП P. incarnata проводилась в окрестностях с. Черкех Таттинского улуса в типологически одинаковых лиственничных сообществах разнотравно-брусничного типа, но находящихся в различных степенях антропогенной нарушенности. По классификации растительности все фитоценозы относятся к ассоциации Aquilegio parviflorae – Laricetum cajanderi Ermakov et al. 2002, большинство которых имеют вторичное происхождение и находятся на различных стадиях дигрессивно-демутационных сукцессий, так как коренные сообщества почти уничтожены в результате интенсивного антропогенного и зоогенного пресса. Всего было рассмотрено 12 ценопопуляций (рис. 1).
Рис. 1. Ряд фитоценозов по степени нарушенности (от более нарушенных к менее нарушенных сообществ)
Изученные ЦП образуют своеобразный экологический градиент от условно благоприятных до сильнонарушенных ценопопуляций. Основное воздействие на ценопопуляции наносит крупный рогатый скот, так как густонаселенные районы Лено-Амгинского междуречья заняты сельским хозяйством.
В последние годы, ослабленные антропогенным влиянием леса этого района, подвергаются нашествиям сибирского шелкопряда и лиственничной чехлоноски. ЦП 5, ЦП 8 и ЦП 11 приурочены к сильно нарушенным фитоценозам. Лиственничные леса ЦП 5 и ЦП 8 были подвержены вспышкам сибирского шелкопряда в 2000 году. ЦП 11 находится на самовосстанавливающейся гари, после пожара в начале 90-х прошлого столетия.
Вторая группа ценопопуляций – это участки в разнотравных лиственничных лесах, которые подвергаются сильному вытаптыванию скотом и людьми. Кроме этого, в ЦП4 в 2010 году произошла вспышка лиственничной чехлоноски, ЦП 3 был подвержен нашествию сибирского шелкопряда.
Третий блок ценопопуляций – это сообщества разнотравных лиственничных лесов, поэтому основной вид воздействия – это умеренный выпас скота.
Остальные ценопопуляции (четвертый и пятый блоки) контрольные, которые, относительно, мало вытаптываются скотом. ЦП 10 находится в активно самовосстанавливающемся лесном участке, после пожара в середине 80-х годов прошлого столетия. ЦП 7 и ЦП 9 это типичные брусничные и бруснично-зеленомошные лиственничные леса с хорошо развитым моховым покровом и с доминированием в травяно-кустарничковом ярусе Vaccinium vitis-idaea, Linneae borealis и Arctous arctostofilos. ЦП 12 находилась в среднеувлажненном разнотравном лиственничном лесу, с довольно богатым видовым разнообразием.
Результаты исследования и их обсуждение
Основными задачами популяционных исследований являются анализ структуры популяций, как возрастных, размерных и виталитетных распределений особей в их составе.
Изучение онтогенетической структуры показало, что вид характеризуется нормальной неполночленной онтогенетической структурой. Исходя из биологических особенностей вида Pyrola incarnatа (для вида характерно вегетативное размножение, нередок процесс омоложения до ювенильного возраста) базовый спектр имеет двувершинную, бимодальную структуру с максимумами особей в виргинильном и субсенильном возрастных состояниях, что показывает вегетативную подвижность объекта. Низкие показатели генеративных состояний показывают что, часть особей виргинильного состояния, минуя генеративное состояние, сразу переходят в субсенильное. Расширение спектра в ювенильном состоянии показывает что в ходе онтогенеза Pyrola incarnatа происходит омоложение до ювенильного состояния. Из таблицы видно что, в малонарушенных фитоценозах (ЦП 6, ЦП 7, ЦП 9 и ЦП 11) доля прегенеративных особей относительно низкая, а постгенеративных высокая. В сильнонарушенных фитоценозах эта картина меняется и доля участия молодых повышается до 92,64 %.
Популяционные показатели и виталитет ценопопуляции Pyrola incarnata
|
ЦП |
Годы |
Возрастная структура |
Плотность |
Классы виталитета |
IVC |
Q |
IQ |
Виталитетный тип |
||||
|
j-v |
g1-g3 |
ss-s |
a |
b |
c |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
|
1 |
2010 |
88,84 |
1,361 |
9,79 |
183,75 |
0,12 |
0,72 |
0,16 |
1,03 |
0,42 |
2,6 |
Процветающая |
|
2011 |
83,03 |
0,45 |
16,52 |
164,02 |
0,04 |
0,68 |
0,28 |
1,09 |
0,36 |
1,3 |
Процветающая |
|
|
2 |
2010 |
74,59 |
0,55 |
24,32 |
91,25 |
0,36 |
0,64 |
0 |
0,87 |
0,5 |
Процветающая |
|
|
2011 |
87,72 |
0,17 |
12,11 |
71,8 |
0 |
0,92 |
0,08 |
0,93 |
0,46 |
5,75 |
Процветающая |
|
|
3 |
2010 |
74,85 |
2,38 |
22,77 |
252,5 |
0,56 |
0,40 |
0,04 |
0,79 |
0,48 |
12 |
Процветающая |
|
2011 |
50,92 |
4,91 |
44,17 |
163,7 |
0,2 |
0,4 |
0,4 |
1,14 |
0,3 |
0,75 |
Депрессивная |
|
|
4 |
2010 |
92,64 |
4,33 |
3,03 |
115,5 |
0,04 |
0,08 |
0,88 |
1,26 |
0,06 |
0,07 |
Депрессивная |
|
2011 |
79,77 |
7,30 |
12,92 |
138 |
0 |
0,32 |
0,68 |
1,17 |
0,16 |
0,2 |
Депрессивная |
|
|
5 |
2010 |
66,66 |
22,09 |
11,24 |
258 |
0,04 |
0,40 |
0,56 |
1,19 |
0,22 |
0,4 |
Депрессивная |
|
2011 |
79,4 |
0,08 |
20,52 |
230,3 |
0,15 |
0,7 |
0,15 |
1,03 |
0,42 |
2,8 |
Процветающая |
|
|
6 |
2010 |
56,67 |
4,082 |
39,25 |
212,33 |
0,04 |
0,84 |
0,12 |
0,94 |
0,44 |
3,7 |
Процветающая |
|
2011 |
64,97 |
1,59 |
33,44 |
274 |
0,28 |
0,44 |
0,28 |
0,96 |
0,36 |
1,3 |
Процветающая |
|
|
7 |
2010 |
56,1 |
10,36 |
33,54 |
246 |
0,2 |
0,56 |
0,24 |
0,98 |
0,38 |
1,6 |
Процветающая |
|
2011 |
74,18 |
0,016 |
25,8 |
231 |
0,2 |
0,52 |
0,28 |
1,03 |
0,36 |
1,3 |
Процветающая |
|
|
8 |
2010 |
76,36 |
4,03 |
19,61 |
359,5 |
0,2 |
0,56 |
0,24 |
1,03 |
0,38 |
1,6 |
Процветающая |
|
2011 |
80,13 |
1,08 |
18,8 |
364 |
0,52 |
0,44 |
0,04 |
0,86 |
0,48 |
12 |
Процветающая |
|
|
9 |
2010 |
54,05 |
4,14 |
41,8 |
265,5 |
0,04 |
0,84 |
0,12 |
0,96 |
0,44 |
3,7 |
Процветающая |
|
2011 |
68,82 |
2,43 |
28,74 |
197 |
0,16 |
0,8 |
0,04 |
0,94 |
0,48 |
12 |
Процветающая |
|
|
10 |
2010 |
72,92 |
17,19 |
9,89 |
128 |
0,12 |
0,68 |
0,2 |
1,08 |
0,4 |
2 |
Процветающая |
|
2011 |
80,16 |
0,01 |
19,83 |
121 |
0 |
0,56 |
0,44 |
0,98 |
0,28 |
0,6 |
Депрессивная |
|
|
11 |
2010 |
60,75 |
2,43 |
36,86 |
82,33 |
0,28 |
0,68 |
0,04 |
0,9 |
0,48 |
12 |
Процветающая |
|
2011 |
91,37 |
0,51 |
8,12 |
129 |
0,36 |
0,64 |
0 |
0,84 |
0,5 |
Процветающая |
||
|
12 |
2010 |
77,99 |
13,84 |
8,18 |
79,5 |
0 |
0,68 |
0,32 |
0,98 |
0,34 |
1,06 |
Процветающая |
|
2011 |
65,1 |
2,68 |
32,21 |
79 |
0,16 |
0,52 |
0,32 |
1,01 |
0,34 |
1,06 |
Процветающая |
|
Плотность грушанки в ценопопуляциях меняется в широких пределах. Наименьшая плотность была в ценопопуляциях 2, 11 и 12, где количество не превышала 100 рамет на 1 м2. Наибольшая плотность была в ЦП 8 359 рамет/м2, которая находится в сильнонарушенном участке. Минимальное значение плотности особей наблюдалось в ЦП 12, которая находится на фоновом участке. Средняя плотность по району исследования составила 184 штук на 1 м2.
По критерию Q депрессивными считаются ценопопуляции 3, 4 и 10. По оценке степени депрессивности наиболее депрессивным является ЦП 4, хотя индекс жизненности (IVC) особей в этих ценопопуляциях достигает своих наивысших показателей (1,14–1,26).Это объясняется тем, что в стрессовых условиях произрастания возрастает генеративное усилие растений. По степени процветания наименьшие показатели отмечены в ЦП 12, в которой IQ (степень отклонения Q) близка к единице, к равновесному типу.
Для выживания в антропогенно-нарушенных лесах Лено-Амгинского междуречья P. incarnata выработал комбинированный защитно-стрессовый тип онтогенетической стратегии, т.е. при нарастании стресса происходит сначала усиление, а затем ослабление координации развития растений. По системе оценки эколого-фитоценотических стратегий Раменского – Грайма грушанка красная обладает смешанным типом жизненной стратегии – конкурентно-стресс-толерантной (SC). В нормальных условиях грушанка ведет себя как патиент (S – стратег), так как в типичных малонарушенных лиственничниках она подавляется видами-виолентами (брусника, арктоус). При любых нарушениях экологических условий (рубка, вытаптывание, выпас и т.д.) происходят сильные изменения условий произрастания (степень увлажнения и освещенности), грушанка проявляет себя как виолент, заселяя новые участки.
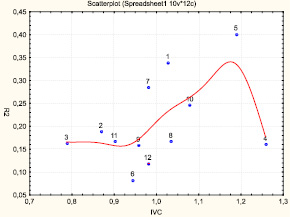
Рис. 2. Тренд онтогенетической стратегии популяции Pyrola incarnata. Номера соответствуют номерам ценопопуляций. По оси абсцисс – индекс виталитета ценопопуляции (IVC), по оси ординат – морфологическая целостность
Заключение
В результате проведенных исследований, ценопопуляций грушанки красной на лиственничных лесах Лено-Амгинского междуречья, можно заключить, что жизненное состояние ценопопуляций может быть оценено как удовлетворительное. Однако в районе исследования с каждым годом усиливается антропогенная нагрузка (усиление выпаса и вытаптывания, развитие туризма и сельского хозяйства др.), что ухудшает эколого-фитоценотические условия обитания. Грушанка красная способна переносить некоторую нагрузку воздействия, даже усиливает свою жизненность, но после этого ослабляет свои позиции.
Библиографическая ссылка
Никифорова А.А. Ценопопуляции Pyrola incarnata нарушенных лесов Лено-Амгинского междуречья (центральная якутия) // Успехи современного естествознания. 2012. № 11-1. С. 47-50;URL: https://natural-sciences.ru/en/article/view?id=31024 (дата обращения: 16.07.2026).