


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
FEATURES OF ANATOMICAL STRUCTURE OF REED MACE IN CONDITIONS OF LEAD OVERLOAD

Высшая водная растительность (ВВР) – один из основных компонентов гидробиоценозов, составляющих локальный природный комплекс. Выполняя многофункциональную роль в гидроэкосистемах, ВВР выступает в итоге как важный фактор регулирования структуры и функций сопутствующих гидробионтов, а также качества воды. При этом степень ее активности специфична биологическим особенностям вида растений, периодам вегетации, адаптационным возможностям, интенсивности антропогенного воздействия. Из токсикантов, попадающих в водоемы, серьезную экологическую проблему представляют тяжелые металлы. Выделена группа наиболее опасных для водных организмов металлов-токсикантов, в которую входят свинец, кадмий, медь, и др.
Свинец – тяжелый металл с уникальными физическими и химическими свойствами, что обуславливает его широкое использование в хозяйственной деятельности человека с древних времен и по настоящее время. Вследствие этого, а также из-за отсутствия свойства биодеградации, его накопление в окружающей среде представляет серьезную экологическую проблему. Свинцовому загрязнению подвержены как наземные, так и водные экосистемы, испытывающие негативные эффекты данного токсиканта в отношении живых организмов разного уровня организации, [8], в том числе высшей водной растительности.
Интегральными показателями состояния гидрофитов в условиях изменения среды обитания являются анатомо-физиолого-продукционные процессы, специфика которых определяется особенностями обитания растения. Эти системы очень лабильны, позволяют быстро перестраивать направленность процессов метаболизма в условиях изменения среды обитания, в частности антропогенной нагрузки. Эта способность может быть использована при определении экологической пластичности растений в условиях нагрузки по токсикантам с различными механизмами действия.
Однако аутэкологические механизмы регуляции адаптационных возможностей водных растений к неблагоприятным условиям среды обитания остаются недостаточно изученными. В этой связи исследование их респонс-реакций на нагрузку по токсикантам, в том числе свинцу, весьма актуально.
Целью данной работы является исследование анатомических особенностей индикаторного гелофита – рогоза узколистного (Typha angustifolia L.) в условиях нагрузки по свинцу.
Материал и методы исследований
Один из наиболее распространенных и популярных способов изучения влияния химических соединений на экосистемы – использование экспериментальных водоемов. Возможность имитации (воспроизведения) разнообразных условий природной среды в лабораторных исследованиях на водных объектах наглядно продемонстрирована в ряде работ, в частности [7].
Исследования проводили в условиях экспериментальных водоемов, включающих природную воду объемом 30 литров с сопутствующими гидробионтами, грунт, куртины представителя ВВР – рогоза узколистного, привезенных из озера Средний Кабан. Оно расположено на территории г. Казани республики Татарстан РФ. При выборе объема экспериментальных водоемов руководствовались методическими данными Тсиртсиса [10], согласно которым объем 30 л позволяет получать удовлетворительную воспроизводимость результатов и хорошее совпадение экспериментальных результатов с расчетами на модели. Моделировали два типа биотопов – заросший (с рогозом узколистным) и открытый (без него). Подготовленные водоемы располагали на специальной площадке научно-исследовательского института.
В качестве загрязнителя использовали свинец в форме соли Pb(CH3COO)2 в концентрации 0,25 мг/л (по свинцу). В экспериментальные водоемы соль вносили однократно через две недели после постановки эксперимента, когда система стабилизировалась, растения прижились (начало июня). Опыты проводили в течение июня – июля в трехкратной повторности. Осуществляли контроль за процессами формирования анатомического строения водных и почвенных корней рогоза.
Анатомические исследования проводили по методике [3]. Поперечные срезы у почвенных корней делали на расстоянии 3 и 6 см от апекса роста, у водных – 6 см. Анатомические исследования у водных корней на расстоянии 3 см от точки роста не осуществляли из-за сложности выполнения поперечных срезов с объекта малого диаметра от руки с помощью безопасного лезвия. Выбор расстояний от точки роста придаточных корней был произвольным. Анализ поперечных срезов проводили в десятикратной повторности, затем рассчитывая стандартное отклонение в программе Excel.
Результаты исследования и их обсуждение
Особенности анатомо-морфологического строения двух типов придаточных корней рогоза узколистного в условиях нагрузки по свинцу
Немаловажную роль в регуляции толерантности макрофита играют особенности строения его подземной части. Она состоит из толстых корневищ, выполняющих роль запасающего органа, и придаточных корней – водных и почвенных (в зависимости от того, в какой среде они развиваются), основной функцией которых является поглощение из внешней среды растворенных питательных веществ и воды [1]. Отличительной чертой анатомического строения является наличие хорошо развитой аэренхимы, которая пронизывает все органы растения и обеспечивает лучший газообмен подземным органам [1, 9].
У макрофитов могут развиваться разные виды придаточных корней (водные, почвенные и стеблевые), сходные по своей этиологии (корневищные), но отличающиеся по количественно-анатомическим признакам и направленностью метаболических признаков.
Ранее нами было установлено, что ответной реакцией рогоза узколистного на нагрузку по азоту является изменение степени аэрации (доли площади воздушных полостей от общей площади поперечного среза) у водных и почвенных придаточных корней [4].
В данной работе ставилась задача: проверить является ли выявленная закономерность универсальной и проявляется при действии загрязнителей с разными механизмами действия.
Контроль
Тип придаточного корня.
В условиях наших экспериментов отмечено, что в плагиотропной зоне корневища располагаются более тонкие ветвящиеся водные корни (через две недели после постановки эксперимента диаметр среза на расстоянии 6 см от апекса роста составил 0,9 ± 0,08 мм), а в ортотропной – относительно толстые (1,3 ± 0,09 мм, соответственно), мало ветвящиеся почвенные. На поперечном срезе придаточных корней в первичной коре между экзодермой и эндодермой расположены тонкостенные клетки паренхимы. В мезодерме отмечены воздухоносные полости – каналы, ограниченные друг от друга несколькими слоями паренхимных клеток. Вглубь за эндодермой на поперечном срезе двух типов придаточных корней расположен осевой цилиндр с проводящими элементами. При этом выявлено, что водные корни более аэрированы по сравнению с почвенными. Так, у водных корней доля площади воздухоносных полостей от общей площади поперечного среза составила 37,2 и 7,2 у почвенных (на расстоянии 6 см от апекса роста через две недели после постановки эксперимента) (табл. 1, 2, рисунок). Вероятно, это связано с особенностями их расположения на корневище и протекания в них метаболических процессов.
а)  б)
б) 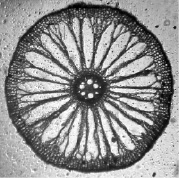
водные корни: а) контроль; б) свинец 0,25 мг/л
а) 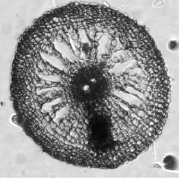 б)
б) 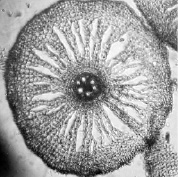
почвенные корни: а) контроль; б) свинец 0,25 мг/л
Фото срезов корней рогоза 6 см от апекса (июль)
Таблица 1
Доля площади (Д, %) воздухоносных полостей (S в.п.) от общей площади поперечного среза (S ср.) почвенных корней Typha angustifolia L. в вариантах опыта (6 см от апекса)
|
Тип |
Почвенные корни |
|||||
|
Дата |
июнь |
июль |
||||
|
Вариант |
Sср (мм2) |
Sв.п (мм2) |
Д, % |
Sср (мм2) |
Sв.п (мм2) |
Д, % |
|
Контроль |
1,53 ± 0,05 |
0,110 ± 0,004 |
7,2 |
1,07 ± 0,07 |
0,16 ± 0,02 |
14,8 |
|
Свинец |
1,519 ± 0,100 |
0,196 ± 0,014 |
12,9 |
1,197 ± 0,130 |
0,238 ± 0,018 |
19,9 |
Таблица 2
Доля площади воздухоносных полостей (Д, %) от общей площади поперечного среза (S ср.) водных корней Typha angustifolia L. в вариантах опыта (6 см от апекса)
|
Тип |
Водные корни |
|||||
|
Дата |
июнь |
июль |
||||
|
Вариант |
Sср (мм2) |
Sв.п (мм2) |
Д, % |
Sср (мм2) |
Sв.п (мм2) |
Д, % |
|
Контроль |
0,90 ± 0,08 |
0,34 ± 0,04 |
37,2 |
0,48 ± 0,03 |
0,16 ± 0,02 |
33,2 |
|
Свинец |
0,605 ± 0,052 |
0,144 ± 0,009 |
23,8 |
0,431 ± 0,048 |
0,065 ± 0,005 |
15,1 |
Возраст растительных тканей
Скорость образования воздухоносных полостей у почвенных корней зависит от возраста паренхимных клеток: чем старше ткань, тем интенсивнее в ней происходят процессы разрушения (в контрольных вариантах на расстоянии 3 см от апекса роста доля воздухоносных полостей на поперечном срезе составила 0 % (табл. 3), тогда как на расстоянии 6 см – 7,2 (табл. 1).
Таблица 3
Доля площади воздушных полостей (S в.п.) от общей площади поперечного среза (S ср.) почвенных корней (3 см от апекса)
|
Вариант (6.06.07) |
Sср (мм2) |
Sв.п (мм2) |
Д % |
|
Контроль |
1,02 ± 0,06 |
- |
0 |
|
Свинец |
1,16 ± 0,09 |
- |
0 |
Количественно-анатомические признаки в условиях наших экспериментов различались у корней в сезонной динамике. Так, у почвенных корней доля площади воздухоносных полостей от общей площади поперечного среза в июне составила 7,2, в июле – 14,8 % соответственно; а у водных корней – 37,8 и 33,3 % соответственно (табл. 1, 2).
Представляло интерес исследование степени воздействия свинца на количественно-анатомические характеристики придаточных корней.
Свинец. Выявленные изменения в анатомическом строении (доля площади воздухоносных полостей от общей площади поперечного среза) у двух типов придаточных корней рогоза в условиях нагрузки по свинцу, также как и в контроле, зависели от типа корня и возраста растительных тканей.
Тип придаточного корня
Почвенные корни. При воздействии уксуснокислого свинца через две недели после постановки эксперимента отмечено повышение степени аэрации корней (увеличение объемов воздухоносных полостей до 12,9 % от общей площади поперечного среза, в контроле – 7,2 %) за счет разрушения тонкостенных паренхимных клеток (табл. 1). Это способствует повышению их энергетического статуса. Как известно, в период адаптации в растении происходят дополнительные энергетические затраты [2], прежде всего за счет усиления поглощения кислорода [6]. У высшей водной растительности часть кислорода образованного в процессе фотосинтеза накапливается в своеобразных резервуарах – в воздушных полостях, который в дальнейшем используется в процессе ее жизнедеятельности [5].
Водные корни. На поперечном срезе водных корней отмечен противоположный эффект – снижение скорости межклеточных разрушений в паренхиме. В контроле площадь воздухоносных полостей через две недели после постановки эксперимента составила 37,2 % от общей площади поперечного среза; в варианте со свинцом 23,82 %, соответственно (табл. 2).
Подобные исследования, проведенные уже через пять недель после внесения одноразовой нагрузки (июль), показали, что количественные показатели доли воздухоносных полостей от общей площади поперечного среза у двух типов придаточных корней изменялись в сторону повышения у почвенных корней и уменьшения у водных (табл. 1, 2, рисунок).
Возраст растительных тканей
Чем старше ткань, тем интенсивнее она подвергается воздействию возмущающего фактора. В варианте со свинцом у почвенных корней на расстоянии 3 см от апекса роста разрушения клеток паренхимы не обнаружено, тогда как на расстоянии 6 см – 12,9 % (табл. 1, 3).
Анализ вышеизложенных данных (по воздействию свинца) свидетельствует о разнонаправленности изучаемых ответных реакций двух типов придаточных корней рогоза узколистного на нагрузку по свинцу: увеличение степени аэрации у почвенных и снижение – у водных по отношению к контролю. Это достигается за счет регулирования интенсивности разрушения паренхимных клеток.. Данное перераспределение позволяет адаптироваться к неблагоприятным условиям среды обитания водным корням, непосредственно контактирующих с растворенными соединениями.
Изменения количественно-анатомических признаков у придаточных корней корневища рогоза узколистного в ответ на вариации минерального питания, по мнению [11], можно рассматривать как механизм поддержания функционирования корней в изменяющихся условиях среды обитания.
Выводы
1. Выявлено перераспределение интенсивности образования воздухоносных полостей, а, следовательно, запасов кислорода, за счет разрушения паренхимных клеток с водных придаточных корней, непосредственно контактирующих с растворенным в воде соединением свинца, на почвенные.
2. Противоположная направленность перестроечных процессов в анатомическом строении двух типов придаточных корней гелофита на нагрузку по уксуснокислому свинцу определяется их разными адаптационными возможностями и условиями обитания.
Работа выполнена при финансовой поддержке Министерства образования и науки РФ в рамках ФЦП «Научные и научно-педагогические кадры инновационной России» (Государственное соглашение № 14.B37.21.0180).
Библиографическая ссылка
Ратушняк А.Ю., Абрамова К.И., Богачев М.И., Полуянова В.И., Андреева М.Г., Чахирев И.В., Ратушняк А.А. ОСОБЕННОСТИ АНАТОМИЧЕСКОЙ СТРУКТУРЫ РОГОЗА УЗКОЛИСТНОГО В УСЛОВИЯХ НАГРУЗКИ ПО СВИНЦУ // Успехи современного естествознания. 2012. № 11-2. С. 38-42;URL: https://natural-sciences.ru/en/article/view?id=31164 (дата обращения: 14.07.2026).