


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
SPECIFIC OF JOINT DWELLING LARVAL POPULATIONS OF RANA ARVALIS and RANA TEMPORARIA

Популяции двух видов Rana arvalis и Rana temporaria населяют одинаковые биотопы и используют для размножения одни и те же водоемы, т.е. характеризуются определенной степенью синтопии. Использование разными видами идентичных участков микроместообитания возможно, благодаря морфологическим или поведенческим смещениям признаков. О наземном и водном периоде жизненного цикла в научной литературе накоплен достаточный материал, установлены экологические ниши, указывающие на отсутствие конкуренции и комплекс факторов, гарантирующих репродуктивную изоляцию. Наиболее полно данные по этому вопросу представлены А.С. Северцовым и др. [5]. Тем не менее, если видоспецифичность наземного периода цикла изучена достаточно, то по водному этапу (эмбриональное и личиночное развитие) остается много неясного. Экспериментальные данные не всегда способны объяснить то, что происходит в естественных условиях, а многообразие естественных условий и отклик на них популяции не позволяет делать однозначные выводы. В задачу данной работы входит определение экологических ниш личинок двух видов бурых лягушек.
Материалы и методы исследования
В связи с множеством разнообразных факторов влияющих на ход икрометания, скорость эмбрионального и личиночного развития, весьма трудно оценить степень видоспецифичности двух видов на разных нерестилищах. По этой причине на протяжении 4 лет (2009–2012 г.) проводились исследования экологических особенностей R. arvalis и R. temporaria на общем нерестилище в Нижнесергинском районе Свердловской области. В этом случае обеспеченность пищей и пресс хищников, в целом емкость местообитаний, следует признать константными из года в год, и исключить их из числа факторов, вызывающих многолетнюю динамику численности конкретной популяции.
Для наблюдения был выбран водоем 1, образовавшийся в песчаном карьере, расположенном в 500 м от реки Серга и находящемся выше уровня реки на 40 м. В течение всего периода личиночного развития хорошо просматривается дно водоема и отсутствует угроза его пересыхания. В весенний период площадь водного зеркала составляет около 3 тыс. м2. Максимальная глубина – 1,5 м, средняя глубина – 0,3–0,5 м. Большая часть береговой линии и дна – пологие.
В 2012 г. под наблюдение был взят нерест травяной лягушки в водоеме 2 со сходными условиями, в 15 м от реки на левом берегу. Однако, за период развития этот водоем пересыхал до небольших луж, вновь наполнялся дождевой водой и зарос злаками, в результате завершение личиночного развития не было зарегистрировано.
На водоемах через каждые 2 дня велся учет появления новых кладок, времени выхода первых личинок, распределения кладок в водоеме.
Благодаря тому, что дно хорошо просматривалось, раз в неделю, пробы личинок брали, ориентируясь на их месторасположение и поведение. Головастиков фиксировали 70 % этанолом и далее определяли стадию развития, размер тела и видовую принадлежность. Визуально на водоеме первую неделю развития видовая принадлежность определяется легко по цвету (рыжие – травяные и черные – остромордые личинки). У фиксированных личинок классическое определение возможно после 43 стадии развития, когда все ряды ротового аппарата сформировались. На ранних стадиях (39–40) может отсутствовать последний зубной ряд у обоих видов, тогда личинка травяной может быть определена, как остромордая. Поэтому учитывался комплекс признаков (длина хвоста, форма верхнего клюва, наличие третьего ряда верхней губы, расположение третьего ряда нижней губы относительно края губы). Эти признаки были установлены нами в результате обработки значительного объема материала при изучении аномалий ротового аппарата личинок. При этом наличие третьего верхнего губного ряда и клювообразная верхняя пластина достаточно редко, но встречается у остромордой лягушки и может отсутствовать у травяной. Наиболее достоверным признаком является расстояние между нижним третьим рядом и сосочками нижней губы. У личинки остромордой лягушки они расположены близко, у травяной есть пространство и даже просматриваются наметки четвертого ряда.
Результаты исследования и их обсуждение
По многочисленным данным R. temporaria приступает к икрометанию на неделю раньше R. arvalis. На урбанизированных территориях оба вида приходят на нерест одновременно [1, 2]. На исследуемой территории, первые кладки остромордой лягушки появлялись одновременно с кладками травяной лягушки (таблица). Это связано с тем, что оба вида зимуют рядом с водоемом, который прогревается значительно раньше тех, что расположены вдоль р. Серги.
Сравнение двух видов в период нереста
|
Год |
2009 |
2010 |
2011 |
2012 |
||||
|
Вид Rana |
tem. |
arv |
tem. |
arv. |
tem. |
arv. |
tem. |
arv. |
|
Начало икрометания |
07.05 |
01.05 |
02.05 |
30.04 |
21.04 |
|||
|
Продолжительность нереста |
5 |
12 |
5 |
10 |
3 |
10 |
3 |
16 |
|
Начало выхода из икры |
– |
06.05 |
10.05 |
01.05 |
||||
|
Кол-во кладок в водоеме / % |
– |
128/22 |
453/78 |
71/18 |
332/82 |
35/6 |
579/94 |
|
Вышедшие личинки R. temporaria, по нашим наблюдениям, переходят в несколько крупных скоплений, которые держатся более 13–18 суток. Не проследили, к сожалению, что стало со студенистыми оболочками икры, но интерес представляет тот факт, что после исчезновения оболочек личинки продолжали держаться вместе. Они располагались на выступающей поверхности дна или камнях, и за неделю в водоеме 2 переместились на 1,5 м. В 2012 году 10 мая численность одного из скоплений на 1 водоеме по приблизительным подсчетам составляла 12 тыс. особей. В это же время отмечены крупные личинки R. temporaria в толще воды.
Головастики остромордой лягушки по окончании эмбрионального развития около суток остаются на месте, присосавшись к подводным предметам, затем рассеиваются по водоему. Распределение их на дне носит случайный характер, В первые две недели они держатся на глубине до 10 см и большая часть их сосредотачивается у самой кромки воды. Скопления по 4–5 особей могут образовываться у подводных объектов (растения, камни). Оболочки кладок обрастают водорослями и примерно через неделю распадаются окончательно.
В исследуемой популяции в 2012 году при одновременном начале нереста и малом количестве травяной лягушки в 1 водоеме, наблюдался ускоренный рост личинок R. temporaria по сравнению с R. arvalis и этим же видом в водоеме 2 (рисунок.) С началом выхода из водоема (14 июня) первых сеголеток R. temporaria средние размеры остромордой лягушки превосходят оставшихся в водоеме травяных. Кроме того, хорошо видно, как меняется коэффициент вариации размеров тела личинок R. temporaria и R. arvalis (рисунок).
Последние кладки остромордой лягушки отмечены 7 мая, а коэффициент вариации к середине периода развития вырос с 15 до 18,5 %. У травяной лягушки при коротком периоде икрометания можно ожидать меньшую вариабельность. Но, 10 мая (6 суток) коэффициент вариации составил 21 %, а через неделю, он понизился до 12 %. На фоне увеличения среднего размера тела личинок снижение вариации указывает на исчезновение мелких особей. Проба личинок минимального возраста 6 дней на водоеме 1 и возраста 9 и 16 дней из водоема 2 были взяты из описанного выше скопления. Увеличение коэффициента вариации на фоне роста размеров указывает на эффект группы, интенсивный рост крупных и ингибирование мелких личинок.
Мелкие личинки, отставшие в развитии, неоплодотворенные икринки служат пищей для крупных особей R. temporaria, что сказывается на их активном росте.
В 2010 году в водоеме 1 на второй и третей неделе личиночного развития наблюдалась массовая смертность личинок. Причины смертности не установлены. У погибших и погибающих личинок была гиперемия на боку брюшной стороны тела или на бедре задней конечности. При обследовании зафиксированных личинок отмечен пустой кишечник. Сделано предположение, что гибель связана с параличом от укусов личинок каких-то хищных насекомых (например, личинок плавунцов ранних стадий развития). Среди погибших в основном крупные личинки травяной и остромордой лягушки. В это же время наблюдалась масса скоплений (по 20–28 особей) личинок травяной лягушки поедающих погибших. Наличие белковой пищи сказалось на высокой скорости роста и развития личинок R. temporaria. Минимальный срок развития составил 34 дня против 37 в 2011 и 2012 гг. В этом же году наблюдается и активный рост личинок остромордой лягушки, которые не отмечены в скоплениях и, вероятно, получали пищу из детрита, обогащенного белками (рисунок).
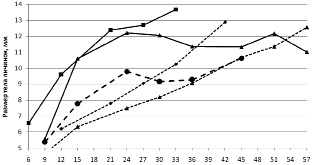
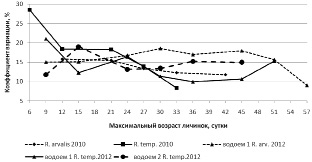
Средний размер и коэффициент вариации размеров тела личинок R. temporaria и R. arvalis в 2010 и 2012 гг.
Таким образом, в 2010 году личинки травяной лягушки получили дополнительную белковую добавку в период раннего и среднего развития, что отразилось на их росте.
Как правило, при опасности личинки травяной лягушки, имея отличную маскировочную окраску, опускаются на дно и не двигались. В 2010 г. они активно реагировали и уплывали на глубину. Личинки остромордой лягушки за все годы наблюдения ведут скрытный образ жизни. Испуганные головастики резко выскакивают из ила и, отплыв на небольшое расстояние, взмучивают грунт и зарываются в него. Под различными укрытиями (камни, доски) также были обнаружены только личинки R. arvalis.
Сопоставляя литературные данные и проведенные наблюдения мы пришли к выводу, что основные различия двух видов заключаются в типе питания, личинки травяной лягушки ближе к плотоядным, а личинки остромордой ближе, к детритофагам. Исходя из вышеизложенного личинкам травяной лягушки свойственен облигатный каинизм, т.е. популяция жертвует одними особями для поддержания жизнеспособности других. Именно этим можно объяснить следующие данные.
R. temporaria откладывает икру сплошными матами. Считается, что таким образом создается микроклимат, а после выхода из икры образуются скопления и действует эффект группы, благодаря которому увеличивается вариабельность по размерам и стадиям развития[5]. В условиях острой конкуренции по темпам онтогенеза гибнут все особи, в фенотипе которых проявляются уклонения онтогенеза, возникшие в период гаструляции и после нее [4]. В пользу каинизма свидетельствуют и крупные кладки до 3,5 тыс. яиц, что в 2–3 раза превышает число икринок у R. arvalis [8].
Ротовой аппарат личинок R. temporaria снабжен мощными роговыми пластинами, верхняя изогнута в виде клюва. У остромордой лягушки роговые пластины с ровным краем и тоньше. Зубные ряды первого вида длинные с хорошо развитыми кератиновыми зубами по сравнению со вторым видом. По данным Е.А. Черданцевой [9], у остромордой лягушки изменчивость структуры ротового поля является по существу модификационной, а у травяной корреляции элементов так высоки, что изменчивость изначально канализирована. Данные Е.М. Писанец и О.В. Ткаченко [3] свидетельствует об опережении головастиками травяной лягушки в формировании практически всех морфологических структур ротового аппарата по сравнению с R. arvalis. Роговой клюв формируется уже на вторые сутки после выхода из икры, у личинок R. arvalis только на четвертые сутки.
Строение ротового аппарата на первом этапе у личинок обоих видов не отличается, на втором этапе происходит увеличение числа вариантов в строении до двух у остромордой лягушки и до восьми у травяной. Это может быть обусловлено отличиями в реакции генотипов на разные условия среды их реализации [3].
Г.С. Сурова и В.Г. Черданцев [6] установили наличие двух дискретных морф яиц (кладки с крупными и мелкими яйцам) у первого вида. Авторы считают, что мелкие яйца принадлежат особям, которые пришли на нерест позднее. Личинки, развивающиеся из яиц мелкой морфы, характеризуются изначально более низкими темпами роста и развития. Однако по мере развития значение этих различий отступает на задний план по сравнению со случайными средовыми влияниями и собственной активностью головастиков. При этом оказывается, что личинки мелкой морфы в большей степени способны к изменению исходных темпов роста и развития.
Для остромордой лягушки характерна высокая вариабельность яиц, как в отдельно взятой кладке, так и между кладками. Увеличению изменчивости размеров тела и скорости развития способствуют еще и растянутые сроки икрометания, 10–13 суток против 3–5 суток у R. temporaria (таблица).
Различия в весе и размере личинок R. arvalis сразу после вылупления незначительны и увеличиваются при переходе головастиков к свободному питанию. В течение двух недель плотность не оказывает влияния на рост и развитие. Эти параметры скорее зависят от особенности самих кладок, и значительно варьирует между кладками [8].
При содержании кладок травяной и остромордой лягушек в лабораторных условиях, первоначальное «загушение плотности» после выхода из икры приводило к быстрому расхождению личинок первого вида по размерам и торможению роста и развития личинок второго вида, которые мало отличались друг от друга в первые 15 суток. Попытки содержания их в сетках на водоеме привели к такому же эффекту [7].
Считается, что пищевая конкуренция отсутствует и во внутривидовых, и в межвидовых отношениях головастиков. Состав микропланктона в толще воды и состав его в кишечниках головастиков из того же водоема совпадает, что свидетельствует об отсутствии избирательности питания [5]. При этом хорошо известно, что удельная скорость роста и скорость развития головастиков, содержавшихся на растительной диете, ниже, чем у содержавшихся на белково-минеральной или смешанной.
Таким образом, если у первого вида увеличение вариабельности размеров связано с механизмом создания увеличенной плотности за счет скопления, при достаточно большом объеме воды, у второго, создается за счет большой вариабельности яиц в кладках и между кладками и растянутого периода икрометания.
В период скопления на кладках у R. temporaria идет отбор на крупных особей, которые растут за счет питания мелкими собратьями. У R. arvalis этот механизм отсутствует и в случае увеличения плотности в естественных условиях рост тормозится у всех особей. Успех развития и размеров первого вида зависит от каннибализма и некрофагии. Это приводит к снижению вариабельности и выходу крупных особей одновременно и в сжатые сроки. При отсутствии такой возможности питание схоже с питанием личинок R. arvalis, при этом внутри и межвидовая конкуренция отсутствует. Личинки остромордой лягушки типичные детритофаги, включая фито и зоофагию за счет мелких объектов питания.
Выявлены различия в поведении личинок. Личинки травяных лягушек, либо активно избегают опасности, либо при мелких размерах маскируются под цвет грунта и обездвижены. Личинки остромордой лягушки приобретают маскировочную окраску во второй половине личиночного развития. При испуге зарываются в грунт или прячутся в укрытия.
Все факты указывают на то, что личинки двух видов занимают совершенно разные экологические ниши в водоеме и действуют разные механизмы регуляции численности, скорости роста и развития.
Библиографическая ссылка
Трубецкая Е.А. ВИДОСПЕЦИФИЧЕСКИЕ ОСОБЕННОСТИ ЛИЧИНОК RANA ARVALIS И RANA TEMPORARIA ПРИ СОВМЕСТНОМ ОБИТАНИИ // Успехи современного естествознания. 2013. № 1. С. 46-50;URL: https://natural-sciences.ru/en/article/view?id=31281 (дата обращения: 14.07.2026).