


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
GEOMETRIC MORPHOMETRICS OF THE MANDIBLE IN CHROMOSOME RACES OF THE PYGMY WOOD MOUSE (SYLVAEMUS URALENSIS PALLAS, 1811): TAXONOMIC AND ECOLOGICAL ASPECTS

Интенсивное развитие методов молекулярного и хромосомного анализа в систематике в начале XXI в. вызвало мощную волну таксономических ревизий и описаний новых видов. Таксономические взаимоотношения между изначально внутривидовыми формами малой лесной мыши (Sylvaemus uralensis Pallas, 1811) также подверглись ревизии. С.В. Межжерин [9, 10] на основе аллозимного анализа и изменчивости морфометрических признаков показал, что выборки малых лесных мышей с Северного Алтая и Памиро-Алая различаются друг от друга и животных из европейской части России и Закавказья, и предложил их считать самостоятельными видами: S. katschenkoi и S. pallipes, соответственно. Он полагал также приемлемой точку зрения Б.А. Кузнецова [8], что на Южном Тянь-Шане и Памиро-Алае обитает S. pallipes, а в районе Алтая, Восточного Казахстана и Северного Тянь-Шаня – S. tokmak [10]. Позже были выявлены различия в кариотипах малых лесных мышей по содержанию прицентромерного гетерохроматина с помощью метода дифференциального С-окрашивания хромосом [1, 7, 11, 15]. А.С. Богдановым были описаны три хромосомные формы S. uralensis, названные по географическому принципу восточно-европейской, южно-европейской и азиатской [1]. Позже было показано, что азиатская хромосомная форма отличается от двух других по кариотипическим и молекулярно-генетическим признакам, поэтому было предложено рассматривать ее в качестве отдельной расы. Восточно-европейская и южно-европейская хромосомные формы были объединены в европейскую расу [2, 4]. Данная схема внутривидовой дифференциации S. uralensis впоследствии была подтверждена результатами нескольких молекулярно-генетических исследований [3, 5, 6, 14]. На основе строения гена цитохрома b мтДНК малой лесной мыши Г.Н. Челомина и Д.М. Атопкин [13] предлагают, однако, рассматривать географические расы малой лесной мыши в качестве самостоятельных таксонов видового ранга. С этой точки зрения только особи западной филогенетической клады могут быть причислены к виду S. uralensis (Pallas, 1811), населяющие ареал от Чехии до Южного Урала. Представители восточной клады, в том числе формы pallipes и katschenkoi, авторы предлагают объединить в составе вида под названием S. tokmak (Severtzov, 1873). Именно эта форма лесной мыши впервые была описана в азиатской части ареала (северная Киргизия).
После описания хромосомной и молекулярной дивергенции названных хромосомных групп необходима ее морфологическая и экологическая верификация. В последние десятилетия разработаны методы геометрической морфометрии [12; 18; 20; 21], позволяющие раздельно анализировать изменчивость размеров и формы объектов и эффективно решить эту задачу.
Исходя из вышесказанного, целью данной работы является изучение степени морфологической дивергенции хромосомных рас и форм (восточно-европейской, южно-европейской и азиатской) малой лесной мыши (Sylvaemus uralensis) методами геометрической морфометрии, используя в качестве объекта анализа форму нижней челюсти. Другой важной задачей было оценить по нескольким морфо-экологическим индексам, полученным по промерам челюсти мышей сравниваемых хромосомных рас, степень их трофической специализации, что можно использовать как дополнительный экологический критерий при оценке возможной видовой самостоятельности.
Материалы и методы исследования
Изученный материал охватывает все три хромосомные группы и представлен выборками из 7-и географических точек (рис. 1). Представители восточно-европейской формы отловлены на территории Урала в трех точках (Свердловская и Оренбургская области, Республика Башкортостан), а южно-европейской формы в Кавказском заповеднике. Выборки из популяций, принадлежащих азиатской расе, взяты в Алтайском заповеднике и в окрестностях г. Каркаралинск и заповеднике «Аксу-Джабаглы» в Казахстане. Использовали в основном коллекционные материалы лаборатории эволюционной экологии ИЭРиЖ УрО РАН и материал, любезно предоставленный к.б.н. Н.Е. Колчевой, а также Зоологическим музеем МГУ. Общий объем материала составил 123 экз. нижнечелюстных ветвей.
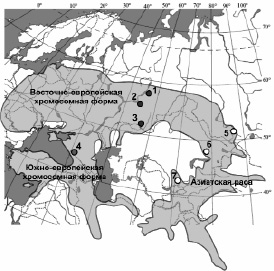
Рис. 1. Карта-схема ареала малой лесной мыши и размещения выборок, относящихся к трем хромосомным группам: восточно-европейской (1–3), южно-европейской (4) формам европейской расы и азиатской расе (5–7). Номера точек – Россия: 1 – Свердловская обл., Шалинский р-н, д. Шигаево (n = 27); 2 – Республика Башкортостан, г. Уфа (n = 12); 3 – Оренбургская обл., Кувандыкский р-н, г. Кувандык (n = 30); 4 – Краснодарский край, Кавказский заповедник, с. Красная Поляна (n = 22); 5 – Республика Алтай, Улаганский р-н, Алтайский заповедник, оз. Телецкое (n = 8); Казахстан: 6 – Карагандинская обл., Каркаралинский р-н, п. Каркаралинск (n = 4), 7 – Южно-Казахстанская обл., Толебийский р-н, заповедник «Аксу-Джабаглы» (n = 10)
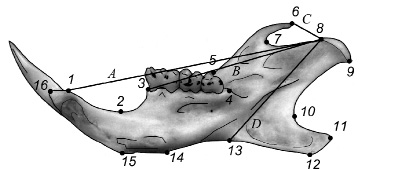
Рис. 2. Размещение меток-ландмарок (1 –16), характеризующих изменчивость формы лингвальной стороны нижней челюсти малой лесной мыши, и промеров, необходимых для вычисления морфо-экологических индексов: A – резцовый, B – молярный, C – темпоральный, D – массетерный
Для изучения морфологической изменчивости выбрали нижнюю челюсть, поскольку она имеет непосредственное отношение к процессам добывания и переработки пищи, которые характеризуют экологические особенности вида. С другой стороны, она является плоским объектом, что важно для корректной оцифровки изображений при использовании методов геометрической морфометрии.
Изображения лингвальной стороны нижних челюстей вводили в компьютер через планшетный сканер Epson V200 с разрешением 1200 dpi. Для характеристики формы нижней челюсти использовали 16 меток-ландмарок (рис. 2), которые размещали на изображениях и оцифровывали с помощью экранного дигитайзера tpsDig2 [19].
Предварительный анализ показал, что различия, связанные с повторной расстановкой меток и повторным получением изображений, оказались статистически недостоверны, что позволяет ими пренебречь. Тем не менее, в дальнейшем использованы усредненные индивидуальные координаты меток по четырем повторам. Для проведения канонического анализа прокрустовых координат применили пакет программ MorphoJ 1.06b [18]. В работе использовали методы ресэмплинга: бутстреп (bootstrap) и перестановочный (permutation) тесты при 10000 повторных реплик.
В ряде работ было показано, что морфо-экологические индексы нижней челюсти, полученные на основе четырех характерных промеров (см. Рис. 2), позволяют изучить конфигурации, характеризующие морфо-функциональные особенности, связанные с различиями в механике обработки пищи грызунами [16; 17]. Тем самым появляется возможность количественно оценить трофическую специализацию внутривидовых форм и видов. Вычисляли четыре индекса [17]: темпорально-резцовый – TI = C/A; темпорально-молярный – TI = C/B; массетерно-резцовый – MI = D/A; массетерно-молярный – MM = D/B. Данные индексы были применены в качестве косвенного экологического критерия видовой самостоятельности.
Результаты исследования и их обсуждение
На рис. 3 в общем морфопространстве вдоль первых двух канонических переменных отображены полигоны изменчивости ординат трех изученных хромосомных групп. Хорошо видно, что азиатская раса существенно отличается от восточно-европейской вдоль первой канонической переменной, но в меньшей степени от южно-европейской формы, полигон изменчивости которой смещен вниз вдоль второй оси. Однако полигон изменчивости последней размещен относительно ближе к восточно-европейской. Полученная картина согласуется с молекулярной филогенией, построенной на основе нуклеотидной последовательности гена цитохрома b мтДНК, а также структуры и формы хромосом. При этом заметный морфологический хиатус проявился лишь между восточно-европейской формой и азиатской расой.
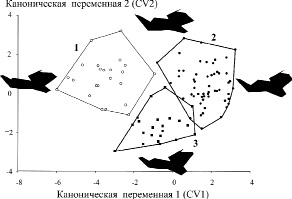
Рис. 3. Канонический анализ изменчивости конфигурации нижней челюсти трех хромосомных групп малой лесной мыши (1 – азиатская раса, 2 – восточно-европейская и 3 – южно-европейская формы) вдоль двух первых осей (CV1, CV2) с учетом теневых моделей челюсти, соответствующих крайним значениям ординат вдоль канонических переменных
На рис. 3 также показаны крайние схематические теневые конфигурации нижней челюсти, характеризующие ее деформации вдоль первых двух канонических осей. Видно, что если у восточно-европейской формы относительно удлинена диастема, вытянут в дорзальном направлении венечный отросток, но укорочен нижний зубной ряд, то у азиатской расы эти черты строения выражены противоположным образом. Конфигурация нижней челюсти у южно-европейской формы из Кавказского заповедника во многом напоминает таковую у восточно-европейской формы, но характеризуется относительно увеличенным угловым отростком и наименьшей шириной мыщелка сочленовного отростка.
Надежность дискриминации всех трех форм сравнительно высока и колеблется в разных сочетаниях от 95,6 до 98,9 %. Проведение проверочного перекрестного теста (cross-validation test), как правило, не приводит к значительному снижению этих величин (от 87,1 до 95,6 %), т.е. указывает на высокий уровень и устойчивость межгрупповых морфологических различий.
Сравнение средних значений морфо-экологических индексов нижней челюсти в выборках двух хромосомных рас, включая южно-европейскую форму, выявило существенный и статистически значимый (p < 0,001) хиатус между европейской и азиатской расами мышей по трем индексам: TI, MI и MM (Табл.). Наибольшие различия проявились по массетерно-резцовому (MI) и массетерно-молярному (MM) индексам (Рис. 4). На рисунке хорошо видно, что выделились две группы, одна из которых объединяет выборки азиатской расы, а другая – европейской (южно-европейская форма близка по индексам к восточно-европейской).
Показатели средних значений морфо-экологических индексов нижней челюсти (± стандартные ошибки) трех хромосомных групп малой лесной мыши
|
Морфо-экологический индекс нижней челюсти |
Европейская раса |
Азиатская раса n = 23 |
|
|
Восточно-европейская форма, n = 68 |
Южно-европейская форма, n = 22 |
||
|
Темпорально-резцовый, TI |
0,157 ± 0,003 |
0,168 ± 0,004 |
0,177 ± 0,007 |
|
Темпорально-молярный, TM |
0,219 ± 0,004 |
0,229 ± 0,006 |
0,238 ± 0,009 |
|
Массетерно-резцовый, MI |
0,551 ± 0,002 |
0,550 ± 0,004 |
0,535 ± 0,003 |
|
Массетерно-молярный, MM |
0,771 ± 0,002 |
0,751 ± 0,005 |
0,721 ± 0,004 |
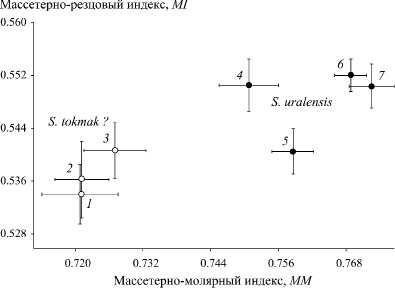
Рис. 4. Сравнение средних значений массетерно-резцового (MI) и массетерно-молярного (MM) индексов нижней челюсти (с учетом величин их стандартных ошибок) в выборках восточно-европейской (1-3) и южно-европейской (4) форм и азиатской (5-7) хромосомной расы малой лесной мыши
Интерпретируя эти различия можно заключить, что если у азиатской расы при обработке пищи в большей степени выражены горизонтальные усилия, связанные с ее перетиранием (индекс TI), то у восточно-европейской, напротив, типичны вертикальные усилия, обеспечивающие дробление и разгрызание объектов питания. Косвенно это указывает на существенную трофическую специализацию обеих рас, а также на их эволюционно-экологическую специфику, включая выполнение ими разных биоценотических функций.
Заключение
Проведенный методами геометрической морфометрии анализ показал, что межрасовые морфологические различия формы нижней челюсти хорошо согласуются с молекулярной и хромосомной гипотезами об относительной филетической близости южно- и восточно-европейских хромосомных форм, но относительно большем генетическом своеобразии представителей европейской и азиатской рас. Морфологическая дивергенция между европейской и азиатской хромосомных расами хорошо выражена. Однако представители южно-европейской хромосомной формы имеют промежуточные черты строения нижней челюсти, хотя и морфологически более близки с восточно-европейской формой. Поэтому нельзя заключить, что между европейской и азиатской расами проявился достаточный хиатус по форме нижней челюсти, позволяющий на индивидуальном уровне безупречно разделить их представителей. Однако следует заметить, что у криптических видов-двойников морфологические различия могут быть выражены даже в меньшей степени. Таким образом, вопрос о видовой самостоятельности азиатской и европейской хромосомных рас эти данные пока решить не позволяют.
В то же время по морфо-экологическим индексам нижней челюсти мышей между азиатской и европейской расами обнаружены резкие различия, указывающие на их трофическую специализацию и, соответственно, экологическую специфику. В данном случае можно говорить о проявлении морфо-функционального хиатуса.
Итак, учитывая аллопатрию рас с очень узкой зоной гибридизации [6], четкое разделение форм по молекулярным и хромосомным характеристикам, а также существенные морфологические различия по форме нижней челюсти и резкий морфологический хиатус по морфо-экологическим индексам, отражающий экологическую специфику форм, есть основания полагать, что представители азиатской расы могут рассматриваться как самостоятельный вид – S. tokmak. Поэтому, не претендуя на окончательный вывод, можно заключить, что полученные новые данные противоречат гипотезе конспецифичности хромосомных рас [5; 6] и скорее подтверждают точку зрения Г.Н. Челоминой и Д.М. Атопкина [13] о видовом статусе S. tokmak.
Исследования выполнены при финансовой поддержке гранта Президента РФ (НШ-2840.2014.4), гранта РФФИ (14-04-31952) и программы Президиума РАН «Живая природа» (12-П-4-1048). Авторы благодарят Зоологический музей МГУ и к.б.н. Н. Е. Колчеву за предоставленный коллекционный материал.
Библиографическая ссылка
Городилова Ю.В., Васильева И.А. ГЕОМЕТРИЧЕСКАЯ МОРФОМЕТРИЯ НИЖНЕЙ ЧЕЛЮСТИ ХРОМОСОМНЫХ РАС МАЛОЙ ЛЕСНОЙ МЫШИ (SYLVAEMUS URALENSIS PALLAS, 1811): ТАКСОНОМИЧЕСКИЙ И ЭКОЛОГИЧЕСКИЙ АСПЕКТЫ // Успехи современного естествознания. 2014. № 11-1. С. 19-24;URL: https://natural-sciences.ru/en/article/view?id=34305 (дата обращения: 27.06.2026).