


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
TO THE KNOWLEDGE OF BIOLOGY OF CISTOGASTER GLOBOSA (FABRICIUS, 1775) (DIPTERA, TACHINIDAE, PHASIINAE), PARASITE OF SOME HEMIPTERA (HETEROPTERA) IN THE MIDDLE-RUSSIAN FOREST-STEPPE

Cistogaster globosa (Fabricius, 1775) – один из семи палеарктических видов рода Cistogaster Latreille, 1829. Первое и единственное сообщение о биологии C. globosa (как Pallasia globosa) в условиях среднерусской лесостепи (Усманский бор, Воронежская область) опубликовано Л.Н. Хицовой и В.Б. Голубом в 1974 г. [Хицова, Голуб, 1974]. В настоящей статье изложены обобщенные результаты многолетнего изучения биологии C. globosa, полученные при обработке коллекционного материала кафедры зоологии и паразитологии Воронежского государственного университета и при проведении наблюдений по биологии вида в природе и лабораторных условиях, с учетом опубликованных нами ранее данных [Хицова, Голуб, 1974].
Материалы и методы исследования
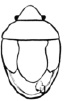
Наши сборы и наблюдения в природе по биологии C. globosa проводились в 1965–2002 гг., преимущественно, в Липецкой, Курской, Воронежской и Тамбовской областях, а также в Горно-Алтайской АО. Кроме того, 1 экземпляр, собранный в Волжско-Камском заповеднике (Республика Татарстан), передан нам В.М. Басовым. Фаунистический материал собирался методами энтомологического кошения по травянистой растительности и выборочной поимки на соцветиях с мая по октябрь. Лабораторные исследования проводились, главным образом, на биостанции Воронежского государственного университета «Веневитиново» (который в настоящее время имеет статус учебно-научного биологического центра). Методика лабораторного эксперимента (рис. 1) описана в работе Л.Н. Хицовой и В.Б. Голуба [1972].

Рис. 1. Копулирующие самец и самка Cistogaster globosa (Fabricius, 1775) в одном садке с хозяином (лабораторный эксперимент)
Результаты исследования и их обсуждение
1. Результаты сборов материала и его определения.
Воронежская область: окр. Воронежа, 19.08.1958, лес Долгий» (1 самец; Скуфьин); 10.09.1958 (1 самка; Скуфьин); лес «Долгий», 2.08.1959 (1 самка; Скуфьин); там же, 23.09.1959 на Sesseli annuum L. (1 самка; Скуфьин); Усманский бор, Веневитино, 17.07.1968, зарастающая гарь (1 самка; Хицова); 26,27,05.1970 (2 самца; Голуб); 29.05.1970 (1 самец, 1 самка; Голуб); 6,8–10.06.1970 (5 самцов, l самка; Голуб); 23–28,30.06.1970 (14 самцов, 12 самок; Голуб); 1,3–5,8.07.1970 (2 самца, 4 самки; Голуб).
Тамбовская область: пос. Знаменка, 19, 22.08.1969, на Matricaria inodora L. (1 самка, 2 самца; Голуб); 22.08.1969 на Аchillea millefolium L. (1 самец, 1 самка; Голуб).
Курская область: Центально-Черноземный заповедник им. проф. Алехина: Стрелецкая степь 31.05.1968, на Fragaria vesca L. (1 самка; Сидорова); 31.05.1968, на Euphorbia virgata Wal. et Rit. (1 самец; Хицова); 14.07.1968, опушка леса, на Matricaria inodora L. (1 самец; Сидорова); 16.07.1968, на Anthriscus sylvestris Hof. (1 самец; Сидорова); 17.07.1968 на Matricaria inodora L. (1 самец; Сидорова); 18.07.1968, склон балки (1 самка; Сидорова); 22.08.1968, на Anthriscus sylvestris Hof. (1 самец; Сидорова); косимая степь; 5,14.07.2002 (2 самца; Лясковская); окр. села Сосновка (вблизи Горшечного); 17.07.1980 (1 самец, 1 самка; Лобанов).
Липецкая область: Галичья гора; 25.05.1965 (1 самец; Кузнецова); 25.08.1965, на Matricaria inodora L. (2 самца; Кузнецова); 2,28.08.1965 (2 самки; Кузнецова); 6.09.1965 (1 самец; Кузнецова); Воргол, опушка леса, 12.07.1970 (1 самец; студ. сборы); Плющань, пойма Дона, 28.07.1971 (1 самка; Кузнецова).
Республика Татарстан: Волжско-Камский государственный заповедник, Сараловский участок, 26.07.2005 (1 самка; Басов).
Горно-Алтайская АО: пос. Артыбаш, западный берег Телецкого озера, в ассоциации с кровохлебкой (Sanguisorba sp.); 1 декада августа 1977 г. (1 самка; Хицова).
2. Биология Cistogaster globosa.
Преферентные биотопы C. globosa – хорошо инсолируемые поляны, опушки и просеки в лиственном или смешанном лесу, ксерофитные участки пойменных лугов с хорошо развитым травянистым покровом, с доминированием представителей семейств зонтичных и сложноцветных. Судя по имеющимся сообщениям и собственным наблюдениям, C. globosa охотно посещает растения родов Achillea, Anthriscus, Matricaria, Euphorbia и др. В мае отдельные экземпляры C. globosa отлавливались с соцветий Fragaria vesca L.
В Центрально-Черноземных областях России и на Украине [Белановский, 1951] C. globosa появляется в начале мая и середине июня, последние поимки одиночных особей (самок) приходятся на август и сентябрь. В Ленинградской обл. [Штакельберт, 1962] самцы и самки C. globosa [как Pallasia globosa] отмечены с июня по август, наибольшее количество собрано в июле-августе. В Армении и Азербайджане самцы найдены в июне и июле [Рихтер, 1971]. В Горно-Алтайской автономной области (пос. Артыбаш) самка, с полностью сформировавшимися яйцами поймана Л.Н. Хицовой в I декаде августа 1972 года.
C. globosa, начиная с тридцатых годов ХХ века, отмечался многими авторами как паразит Aelia acuminatа (L.) и Ae. rostrata Boh. Михалком [Michalk, 1940], Дюпюи [Dupuis, 1963]; Драбер-Монько [Monko, 1957]; Гертингом [Herting, 1971], эти же виды клопов и Ae. sibirica Reuter указаны в 2002 в качестве хозяев C. globosa для Великобритании [Paper & Smith, 2002].
Значительно меньше сведений о паразитировании C. globosa F. на клопе Neotiglossa leporine (H.-Sch.) [Dupuis, 1963]. В отечественной литературе этот вид в качестве хозяина C. globosa до публикации Л.Н. Хицовой и В.Б. Голуба [1974] не отмечался. Cведения о развитии, пищевых связях и фенологии C. globosa также малочисленны.
По нашим наблюдениям в Усманском бору, клопы Ae. acuminatа и N. leporina являются для C. globosa равноценными хозяевами (по крайней мере, в годы наблюдений) в связи с таксономической близостью, общностью стаций, заселяемых этими видами клопов, сходством жизненных циклов и симпатрическими ареалами. Наиболее характерные для них биоценозы в Усманском бору располагаются на припойменных террасах реки Усмани. К ним относятся поляны в судубраве, окраины осоковых болот с преобладанием в травянистом покрове овсяницы луговой (Festuca pratensis Huds.), тростника обыкновенного (Phragmites communis Trin.) и мятлика лугового (Рoа pratensis L.). Откладка яиц, питание личинок и взрослых клопов отмечены только на злаках, в частности, на овсянице луговой.
C. globosa развивается в своих хозяевах в двух поколениях. При этом сроки развития обоих поколений паразита сопряжены со сроками развития хозяев, имеющих моновольтинный цикл развития с имагинальной диапаузой. Заражение хозяина и развитие C. globosa представляется следующим образом.
C. globosa уходит на зимовку в фазе личинки II поколения, находясь в организме имаго зимующего хозяина. Так, на опытных участках с Festuca rubra L. в пойме р. Дон (заповедник «Галичья гора», Липецкая область) в мае 1967 год собраны и затем вскрыты 40 перезимовавших имаго Ae. acuminatа, среди которых 10 особей оказались с личинками C. globosa. Во время сборов шла интенсивная копуляция клопов.
C. globosa покидает зараженного хозяина, выталкивая его анальный сегмент, и, попав в почву, окукливается. Хозяин при этом погибает. Оставшиеся в живых незараженные перезимовавшие клопы проходят период дополнительного питания. Закончившие развитие и вылетевшие из пупариев самки C. globosa первого поколения после копуляции в конце мая-начале июня заражают часть перезимовавших клопов, продолжающих питаться. В конце июня-июля появляются завершившие развитие имаго C. globosa, вышедшие из зимовавших клопов. Самки фазиины вскоре после копуляции откладывают яйца на нимф и взрослых клопов нового поколения.
На зараженных в природе и in vitro клопах Ae. acuminatа яйца C. globosa обнаружены на III-V брюшных тергитах с правой и левой сторон симметрично. Микропиле отложенных яиц направлено назад, в сторону вершины брюшка хозяина с небольшим отклонением вправо или влево. Аналогичным было заражение клопа Ae. acuminatа и в лабораторных условиях. При этом количество отложенных яиц на одном экземпляре было большим, однако локализация их оставалась той же, с сохранением тенденции к симметричности. Так, в случае откладки на одного самца Ae. acuminatа 13 яиц, 7 на них размещались слева и 6 справа. Иногда отмечалась полная или частичная накладка яиц друг на друга.
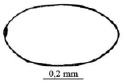

А Б В


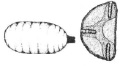
Г Д Е Ж
Рис. 2. Стадии развития Cistogaster globosa (Fabricius, 1775 ): А – расположение яиц на хозяине; Б – форма яйца; В, Г, Д, – ротоглоточные аппараты личинок 1–3 возрастов, Е – форма пупария и конические выступы дыхалец; Ж – заднее дыхальце
Заражение N. leporina имело иной характер. Большая часть яиц откладывалось также на брюшные тергиты, но дисперсно: кроме III–IV сегментов для размещения яиц использовались II и VI сегменты (рис. 2, А).
Яйца C. globosa обнаружены также на второй паре крыльев, в апикальной части, под прикрытием перепоночки надкрыльев (этот вариант откладки яиц отмечен как в природе, так и в лаборатории). Очевидная дисперсность откладки яиц, возможно, связана с габитуальными особенностями хозяев: Ae. acuminatа имеет более продолговатое тело, тогда как N. leporina – более короткое и широкое.
Яйца C. globosa относят к макротипу. Оно имеет выпуклую дорзальную поверхность и плоскую вентральную. Сверху форма яйца овальная или слегка вытянутая (рис. 2, Б). Средняя его длина составляет 562 + 1,97 мк, средняя ширина = 336 + 1,42 мк. Своеобразна и отлична от яиц других фазиин аэропилярная зона (по результатам нашего исследования с помощью сканирующего микроскопа). Она представлена несколькими лопастями, прикрывающими отверстия пластрона. Окраска яиц изменчива: отложенные на брюшные тергиты, как правило, обладают светло-серым, почти белым или слегка буроватым цветом; расположенные открыто – на крыльях, или щитке – темнее. Промежуточная окраска – от темной до светло-бурой – обмечена у яиц, находящихся на одном из надкрыльев под прозрачной перепоночкой другого. Спустя несколько часов после откладки, дорсальная поверхность яйца светлеет, с выходом личинки цвет изменяется до светло-серого, кремового и даже белого.
Длительность эмбрионального развития, начиная от откладки яйца, при температуре 25–30 °С и относительной влажности, близкой к 100 %, составляет 115–140 часов (примерно 5 дней).
Морфологическая характеристика личинок C. globosa дана Нильсеном [Nielsen, 1916]. Мы ограничимся описанием строения ротоглоточных аппаратов личинок младших возрастов. Наиболее значима структура непарного ротоглоточного аппарата личинки I возраста: на переднем крае имеется крупный апикальный клювообразный выступ, позади которого расположена небольшая выемка (рис. 2, В, Г, Д). За выемкой следует волнообразный выступ, переходящий в хорошо выраженное углубление, которое отделяет переднюю непарную часть ротового склерита от задней, расчлененной на дорсальный и вентральный отростки. Дорсальный отросток значительно сужается в дистальному концу, вершина его острая, нижняя сторона слабо скеротизована, за исключением проксимального участка. Вентральный отросток, разделенный на сильно и слабо склеротизованные участки, заметно сужается к своей вершине. Дуговидно изогнутый лабиальный склерит очень тонкий. Личинка I возраста, как и остальных возрастов фазиин, метапнестична, дыхательные отверстия имеют двулопастной характер. Ротоглоточный аппарат личинки III возраста состоит уже из трех отделов: ротовых крючьев, гипостомального и фарингеального склеритов. Задние отростки ротовых крючьев тупые и резко изогнуты (рис. 2, Д).
В строении ротоглоточного аппарата личинки III возраста C. globosa (рис. 2, Д) много общего с таковым личинок старшего возраста видов рода Gymnosoma Meigen, 1803. Отличие заключается в том, что у C. globosa имеются довольно длинный гипостомальный склерит и крупный вентральный выступ. Личиночное развития C. globosa длится около 15 дней.
Пупарий темно-бурого цвета, длиной до 5 мм (рис. 2, Е). На заднем конце расположена пара дыхальцевых выростов (рис. 2, Ж). Дыхальца с тремя спиракулярными щелями (рис. 2, 3). Куколка развивается в течение 13–15 дней, а в целом (в лабораторных условиях) развитие C. globosa, начиная с откладки яйца, длится 35–40 дней.
Заключение
На основании многолетних сборов фазиины C. globosa в среднерусской лесостепи установлено, что вид не многочислен. Биотопически приурочен преимущественно к заселяемым биотопам его хозяев – клопов-щитников Aelia acuminata и Neittiglossa leporina. Сборы в природе и эксперименты по заражению клопов C. globosa позволяют утверждать, что не только виды рода Aelia (в условиях наших наблюдений – Aelia acuminata), но и клоп Neottiglossa leporina активно используется этой фазииной в качестве хозяина. Учитывая несколько разноплановую локализацию макротипических яиц C. globosa на одном и другом хозяине, можно предполагать лишь относительную топическую специализацию в освоении хозяина как субстрата для заражения, возможно, определяемую их габитуальными различиями.
Представляется, что из диагностических признаков (уровень преимагинальных стадий жизненного цикла) достаточно высокую значимость может иметь строение аэропилярной зоны яйца и строение ротоглоточного аппарата личинки I возраста.
Библиографическая ссылка
Хицова Л.Н., Голуб В.Б. К ПОЗНАНИЮ БИОЛОГИИ CISTOGASTER GLOBOSA (FABRICIUS, 1775) (DIPTERA,TACHINIDAE, PHASIINAE), ПАРАЗИТА НЕКОТОРЫХ ПОЛУЖЕСТКОКРЫЛЫХ (HETEROPTERA) В УСЛОВИЯХ СРЕДНЕРУССКОЙ ЛЕСОСТЕПИ // Успехи современного естествознания. 2014. № 12-1. С. 43-46;URL: https://natural-sciences.ru/en/article/view?id=34483 (дата обращения: 13.07.2026).