


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
COPY NUMBER VARIATIONS OF APOPTOSIS-REGULATING GENE AS INDICATOR OF STOMACH TISSUE MALIGNANCY

В настоящее время в мире рак желудка является четвертой по частоте заболеваемости у мужского населения и пятой – среди женского, и второе место в структуре общей летальности от злокачественных новообразований. Среди 45 стран мира Россия занимает 1-е место по уровню смертности от данного заболевания [5].
Трансформация нормальных клеток в раковые связана с накоплением изменений в геноме под действием наследуемых и приобретенных мутаций [10]. Трансформированные клетки получают возможность выживать в организме за счет изменений в апоптозных сигнальных путях, что обеспечивает приобретение неограниченного потенциала к пролиферации и способности к инвазии и метастазированию. Молекулярные изменения, ответственные за приобретение вышеуказанных свойств, могут использоваться в качестве онкомаркеров [1].
Процесс апоптоза можно разделить на три фазы: сигнальную, эффекторную и деградационную. Несмотря на разнообразие инициирующих факторов, выделяются два основных пути передачи сигнала апоптоза: рецептор-зависимый и митохондриальный. Рецептор-зависимый сигнальный путь реализуется через взаимодействие рецептора Fas с адаптером FADD, который взаимодействует с прокаспазой 8. Митохондриальный сигнальный путь реализуется в результате выхода апоптогенных белков из межмембранного пространства митохондрий в цитоплазму клетки вследствие повышения проницаемости наружной мембраны митохондрий, существенную роль в этом играет апоптотический белок Bax. В течение эффекторной фазы происходит активация каспазного каскада. Инициаторные каспазы (– 2, – 8, – 9, – 10, – 12) активируют эффекторные (– 3, – 6, – 7) каспазы, которые в свою очередь непосредственно участвуют в трансформации клетки. В итоге морфологические и биохимические изменения приводят к гибели клетки [3, 4]. Важную роль играет белок p53, который принимает участие в запуске апоптоза путём стимуляции рецепторов смерти, путём взаимодействия с промотором апоптоза – Bax, путём активации p53-зависимого модулятора PUMA (p53 upregulated modulator of apoptosis), который блокирует действие Bcl-2 [9].
Изменение относительного числа копий гена является одним из основных механизмов контроля раковой клеткой ключевых для выживания и малигнизации экспрессии генов. Копийность генов или вариация числа копий генов (сopy number variation, CNV) – вид генетического полиморфизма, результатом которого может явиться снижение или повышение относительного числа копий определенного гена, и, следовательно, пониженная или повышенная экспрессия продукта гена – белка или не кодирующей РНК [8].
Изучена амплификация гена MDM2 при раке желудка, и показано, что увеличение копийности этого гена коррелирует с увеличенной экспрессией белка MDM2 и понижением экспрессии белка P53. Ген MDM2 считается негативным регулятором функции белка Р53. MDM2/P53 путь является важной составной частью канцерогенеза. Только около 20 % случаев рака желудка не связаны с изменением копийности генов MDM2 и P53 [7].
Идентификация генов, которые как сверхэкспрессируется и амплифицируются, так и снижают экспрессию и делетируются очень важна, потому что эти гены могут представлять инициаторную точку генетических изменений.
Цель исследования
Изучить механизмы малигнизации тканей желудка на примере изменения относительной копийности генетических локусов, ответственных за апоптоз.
Материалы и методы исследования
Изучены образцы тканей (опухолевые и условно здоровые), которые были получены в процессе хирургического вмешательства в Ростовском научно-исследовательском онкологическом институте с 2013 по 2014 гг., у 29 пациентов с различным гистологическим типом рака желудка: аденокарцинома G1-G2 (15 пациентов), G3 (5 пациентов), перстневидноклеточный рак (5 пациентов) и смешанного типа (аденокарцинома G3-перстневидноклеточный рак) (4 пациента). Все пациенты, вошедшие в данное исследование, имели ECOG статус от 0 до 2. Для верификации образцов тканей проводилось стандартное патолого-морфологическое исследование с окрашиванием фиксированных срезов гематоксилин-эозином. Биоптаты тканей классифицировали на две группы: опухолевые (малигнизированные) и контрольные (не малигнизированные) образцы.
В дальнейшем геномную ДНК экстрагировали из свежезамороженных биоптатов с использованием лизирующего SDS-содержащего буфера в присутствии протеиназы-К и последующей фенол-хлороформной экстракцией [2]. Концентрацию полученных препаратов ДНК измеряли на флюориметре Qubit 2.0® (Invitrogen, США) с использованием набора Quant-iT™ dsDNA High-Sensitivity (HS) Assay Kit (Invitrogen, США). Для проведения Real-Time qPCR концентрацию образцов ДНК нормализовывали до величины 2 нг/мкл.
Метод определения относительной копийности генетических локусов методом Real-Time qPCR, заключается в одновременной амплификации гена-мишени и референтного гена в опытной и контрольной пробах. Вывод об изменении дозы гена делается на основании соотношения сигналов, продуцируемых амплификатами изучаемой и референсной последовательностей. Прямые и обратные праймеры были разработаны с использованием референтных последовательностей ДНК NCBI GenBank (таблица). Каждые 25 мкл ПЦР-смеси содержали 10 нг геномной ДНК, 0,2 mM dNTP’s, по 100 нМ прямого и обратного праймеров для референтного гена (RNaseP) или гена-мишени, 2,5 mM MgCl2, ПЦР-буфер, 0,05 u/µl ДНК-полимераза Thermus aquaticus («Синтол», Россия), краситель SYBR®Green I (Invitrogen, США). Амплификация каждой пробы осуществлялась в трех повторениях.
Количественная RT-PCR амплификация проводилась с использованием термоциклера Bio-Rad CFX96 (Bio-Rad, USA) в соответствии с инструкциями производителя по следующей программе: 95°C 3 мин., и 40 циклов при 95°C 10 сек, 58°C 30 секунд (чтение оптического сигнала красителя FAM для красителя SYBR-Green) и 72°C 15 секунд. Первичные данные RT-qPCR получали с использованием программного обеспечения Bio-Rad CFX Manager (ver. 2,1). Генетический локус B2M использовали в качестве референтного для нормализации полученных показателей количественной RT-qPCR.
Усредненные данные по каждому генетическому локусу нормировались по усредненному показателю референтного гена B2M для получения величины ΔCt (ΔCt = Ct(исследуемого гена) – Ct(B2M)). Относительную копийность генетического локуса (RQ) рассчитывали по формуле 2-ΔCt. Далее вычисляли медиану RQоп опухолевых образцов и медиану RQк контрольных для каждого генетического локуса и рассчитывали соотношение относительной копийности генов в опухолевой ткани по отношению к нормальной ткани: RQоп / RQк.
Результаты исследования и их обсуждение
У пациентов с аденокарциномой (G1-2) нами были исследованы 10 генетических локусов, 2 из которых были использованы в качестве референтных (ACTB и B2M), и была выявлена статистически не достоверная тенденция к амплификации генетических локусов Casp8 и Mdm2 на 137 и 377 % соответственно и тенденция к снижению копийности генов CASP3 и P53 на 27 и 23 % соответственно (рис. 1). Существенного и статистически достоверного изменения копийности других генетических локусов регуляции апоптоза (BAX, BCL2, CASP8/FADD, CASP3, CASP9, P53) в тканях пациентов с аденокарциномой (G1-2) в ходе данного исследования мы не обнаружили.
У пациентов с аденокарциномой желудка (G3) в рамках настоящего исследования обнаружены статистически достоверные снижения копийности 5 генетических локусов в опухолевой ткани желудка по сравнению с нормальной тканью (рис. 2).
Панель праймеров для определения относительной копийности генов
|
№ п/п |
Наименование |
№ NCBI GenBank |
Хромосомная локализация |
|
1 |
ACTB |
NM_001101.3 |
7p22 |
|
2 |
B2M |
NM_004048.2 |
15q21–q22.2 |
|
3 |
BAX |
NM_138761.3 |
19q13.3–q13.4 |
|
4 |
CASP3 |
NM_004346.3 |
4q34 |
|
5 |
CASP8 |
NM_001080125.1 |
2q33–q34 |
|
6 |
BCL2 |
NM_000633.2 |
18q21.3 |
|
7 |
CASP8/FADD |
NM_003879.5 |
2q33–q34 |
|
8 |
P53 |
NM_000546.5 |
17p13.1 |
|
9 |
CASP9 |
NM_032996.3 |
1p36.21 |
|
10 |
MDM2 |
NM_002392.5 |
12q13–q14 |
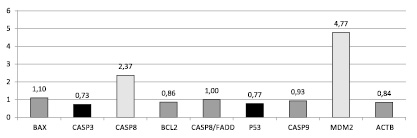
Рис. 1. Соотношение относительной копийности генов в опухолевой и нормальной ткани пациентов с аденокарциномой (G1-2)
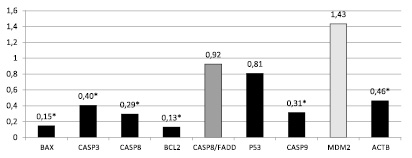
Рис. 2. Соотношение относительной копийности генетических локусов в опухолевой и нормальной ткани пациентов с аденокарциномой (G3) (* – достоверные отличия, p < 0,05)
Относительная копийность была статистически достоверно снижена для порогового уровня p < 0,05 для генов: Bax на 85,5 %, Casp3 на 59,7 %, Casp8 на 70,6 %, Bcl2 на 87,1 % и Casp9 на 68,8 %. Также наблюдалось статистически достоверное снижение копийности гена Actb (housekeeping genes), кодирующего белок β-актин, на 53,7 %. Относительная копийность генетических локусов Bax и Bcl2, кодирующих белки ответственные за активацию и подавление митохондриального апоптоза, изменялись однонаправленно, снижаясь на 85 и 87 % соответственно. Уменьшение относительной копийности затронуло гены, кодирующие как регуляторные (Casp8 и 9), так и эффекторные каспазы (Casp3). Статистически достоверного увеличения относительной копийности генов на данной стадии дифференцировки опухоли не обнаружено, наблюдалась лишь тенденция к амплификации генетического локуса Mdm2 на 43 % и к снижению копийности гена p53 на 19 %.
У пациентов со смешанным типом рака (аденокарцинома G3 + перстневидноклеточный рак) обнаружено статистически достоверное для порогового уровня (p < 0,05) снижение копийности генов Bax на 24 %, Casp8 на 22 % и Casp9 на 19 %. При этом обнаружена тенденция к амплификации генов Bcl2 и Casp8/Fadd на 86 % и 73 % соответственно и тенденция к снижению копийности гена MDM2 на 63 % (рис. 3).
У пациентов с перстневидноклеточным типом рака желудка обнаружено статистически достоверное снижение относительной копийности гена p53 на 46 % (p < 0,01) по сравнению с условно нормальной тканью (рис. 4).
При этом относительная копийность гена Bcl2 статистически достоверно увеличилась на 90 % (p < 0,05). Также обнаружена тенденция к амплификации генов Bax и Casp8/Fadd на 68 % и 75 % соответственно и тенденция к снижению относительной копийности генов Casp9 и Actb на 44 % и 20 % соответственно.
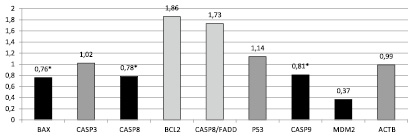
Рис. 3. Соотношение относительной копийности генов в опухолевой и нормальной ткани пациентов со смешанным типом рака (* – достоверные отличия, p < 0,05)
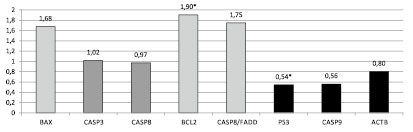
Рис. 4. Соотношение относительной копийности генов в опухолевой и нормальной ткани пациентов с перстневидноклеточным раком (* – достоверные отличия, p < 0,05)
Полученные нами данные свидетельствуют о том, что изменение относительной копийности генов в опухолях разных гистологических типов и на разных стадиях дифференцировки опухоли происходит неодинаково.
В нашем исследовании отмечается факт достоверного повышения (на 90 %) и выраженной тенденции к повышению (на 86 %) относительной копийности гена Bcl2 в опухолевой ткани перстневидноклеточного и смешанного рака (аденокарцинома G3 + перстневидноклеточный) в противоположность со снижением относительной копийности этого же гена, при аденокарциноме G3 (на 87 %) и незначительной тенденции к снижению при аденокарциноме G1-2 (на 14 %). По данным литературы, биологически и этиологически у разновидностей рака желудка имеются различия и в экспрессии онкогенов и в инактивации генов-онкосупрессоров [9], следовательно, отличия могут наблюдаться и в относительной копийности одних и тех же генов, и в опухолевых тканях желудка разного гистотипа. Корреляционный анализ изменения относительной копийности исследуемого паттерна генов в опухолевых тканях показал наличие положительной корреляционной связи между изменениями, обнаруженными у пациентов с аденокарциномой G3 и аденокарциномой G1-2 (r = 0,663), положительной корреляционной связи между изменениями, обнаруженными у пациентов со смешанным типом рака (аденокарцинома G3 + перстневидноклеточный) и перстневидноклеточным раком (r = 0,624), и отрицательной корреляционной связи между изменениями, обнаруженными у пациентов аденокарциномой G3 и смешанным типом рака (r = 0,608). Этот факт демонстрирует, что изменения относительной копийности в генетических локусах, регулирующих апоптоз, при различных гистотипах рака желудка имеют не только количественный, но и качественно разный характер.
Наблюдаемая при высокодифференцированной аденокарциноме (G1-2) тенденция к увеличению копийности гена MDM2 на 377 % сопровождается тенденцией к снижению копийности P53 на 23 %. Схожая тенденция к амплификации генетического локуса MDM2 на 43 % и к снижению копийности гена P53 на 19 % наблюдается и при переходе к умеренно- и низкодифференцированной аденокарциноме (G3). Направление этих изменений, обнаруженное в ходе исследования было ожидаемым, так продукт гена MDM2 является негативным регулятором P53, и подобные изменения описаны для рака желудка [7]. Однако достоверное снижение копийности гена P53 в нашем исследовании наблюдается только в опухолевых тканях желудка у пациентов с перстневидноклеточным раком. А у пациентов со смешанным типом рака копийность гена P53 не изменяется.
Неодинаково изменяется относительная копийность генов, кодирующих регуляторные (Casp8 и 9) и эффекторные каспазы (Casp3), у пациентов с разным гистологическим типом желудка. Так при аденокарциноме (G1-2) имеется тенденция к снижению копийности CASP3 на 27 %, при аденокарциноме (G3) наблюдается достоверное снижение копийности на 59,7 %. А при смешанном и перстневидноклеточном раке копийность этого гена не изменяется. Гены регуляторных каспаз (CASP8 и 9) достоверно снижают свою копийность только у пациентов с аденокарциномой G3 и смешанным типом рака. У пациентов с перстневидноклеточным раком имеется лишь тенденция к снижению копийности CASP9.
Копийность гена Casp8/Fadd статистически достоверно не изменяется, однако у пациентов со смешанным типом (аденокарцинома G3 + перстневидноклеточный рак) и перстневидноклеточным раком имеется тенденция к увеличению копийности этого гена в опухолевой ткани по сравнению с условно нормальной на 73 и 75 % соответственно, что могло бы свидетельствовать о потенциально большей вероятности активации внешнего сигнального пути апоптоза. Интересно, что у пациентов с этими двумя гистологическими типами рака желудка также наблюдается тенденция и достоверное увеличение копийности гена анти-апоптозного белка Bcl2 на 86 % и 90 % соответственно.
Таким образом, полученные данные свидетельствуют о важной роли изменения копийности апоптоз-регулирующих генов BAX, CASP3, CASP8, BCL2, P53 и CASP9 в малигнизации тканей желудка и возможном использовании их относительной копийности в качестве онкомаркеров для разных гистологических типов рака желудка.
Библиографическая ссылка
Кит О.И., Водолажский Д.И., Кутилин Д.С., Малейко М.Л., Двадненко К.В., Енин Я.С., Гудуева Е.Н., Ильченко С.А. ОТНОСИТЕЛЬНАЯ КОПИЙНОСТЬ АПОПТОЗ-РЕГУЛИРУЮЩИХ ГЕНОВ КАК ПОКАЗАТЕЛЬ МАЛИГНИЗАЦИИ ТКАНЕЙ ЖЕЛУДКА // Успехи современного естествознания. 2015. № 3. С. 40-45;URL: https://natural-sciences.ru/en/article/view?id=34733 (дата обращения: 07.06.2026).