


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
DIFFERENTIATION AND SPECIALIZATION OF STRUCTURES IN THE EVOLUTION OF SOME GROUPS OF EMBRYOPHYTES

Дифференциация и специализация были основными модусами структурной эволюции в разных группах высших растений. Они реализовались на морфологическом и анатомическом уровнях. При значительном структурном разнообразии всех крупных таксонов высших растений относительная роль этих преобразований оказалась различной. При модульном типе организации, характерном для всех высших растений, особое эволюционное значение имели признаки, сопряженные с изменением морфогенетических программ и общей программы онтоморфогенеза [7, 8, 11]. В биологической систематике cпецифика любого таксона по сравнению с другими систематическими группами выясняется посредством конструирования его архетипа [4, 6]. При выявлении архетипов таксонов модульных организмов актуален поиск общих особенностей алгоритмов развития, которые проявляются на разных уровнях структурной иерархии. Именно они формируют основу архетипа таксона модульных организмов [7]. С этой точки зрения интересен специальный анализ характера встречаемости структур, обладающих разной степенью дифференциации и специализации, на морфологическом и анатомическом уровнях в крупных таксонах высших растений. В качестве таких значительных по объему и уровню разнообразия групп особенно интересны однодольные и двудольные цветковые растения (классы Magnoliopsida, Liliopsida) и два наиболее крупных по числу видов отдела мохообразных – Bryophyta и Marchantiophyta [7, 9].
Материалы и методы исследования
Для выявления основных закономерностей встречаемости структур с более высоким уровнем дифференциации и специализации изучено их распространение в классах Magnoliopsida, Liliopsida и отделах Bryophyta и Marchantiophyta. У мохообразных специально проанализированы группы, выделяемые в зависимости от характера расположения архегониев. Среди них верхоплодные и бокоплодные мхи (класс Bryopsida, группы Acrocarpae и Pleurocarpae), акрогинные и анакрогинные юнгерманниевые печеночники (класс Jungermanniopsida, группы Acrogynae и Anacrogynae) (таблица).
Оценена встречаемость структур с разной степенью дифференциации и специализации. Выяснен характер распространения структур с предельно сложным строением и максимальным уровнем дифференциации. В совокупности эти характеристики позволяют оценить полноту реализации модусов дифференциации и специализации в структурной эволюции рассмотренных групп.
Результаты исследования и их обсуждение
В классах цветковых растений разная относительная роль модусов дифференциации и специализации проявилась достаточно четко на разных структурных уровнях. Различные варианты специализации и дифференциации выявляются у двудольных растений (класс Magnoliopsida) в архитектуре тела, в строении основных органов и структур, в анатомии осевых органов.
У двудольных четко выражена иерархическая дифференциация корней в корневых системах, жилок в васкулярной системе листьев, часто встречаются листья с прилистниками и цветки с двойным околоцветником (таблица). Представлены вторичные меристемы – камбий и феллоген. Древесные двудольные растения характеризуются значительным разнообразием архитектурных моделей со сложной иерархической дифференциацией осей в кроне. Она характерна, например, для моделей Rauh, Attims, Stone, Aubrevill, Scarrone [10]. Именно у двудольных отмечены соцветия с флоральными единицами высокого уровня псевдоциклической агрегации. Например, у некоторых зонтичных, бобовых, сложноцветных описаны соцветия, соответствующие третьему и четвертому уровням псевдоциклического ряда [2, 3].
Характер распространения структур с разной степенью дифференциации и специализации в некоторых группах высших растений
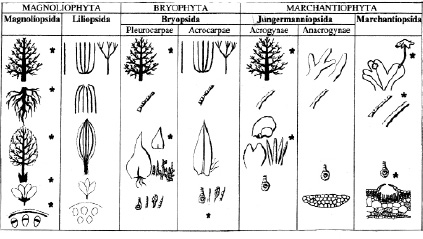
Примечание. * – дифференциация, специализация и иерархия структурных единиц, которая противопоставляется их равноценности и эквивалентности.
Морфогенетические программы однодольных цветковых растений не предполагают сложную иерархическую дифференциацию структуры и элементов разного уровня. Равноценными и эквивалентными являются корни в корневых системах, жилки в васкулярной системе листьев, как правило, не выражены прилистники, редко встречаются цветки с двойным околоцветником (таблица). Можно отметить также относительно меньшую степень анатомической дифференциации осевых органов, отсутствие специализированных латеральных вторичных меристем. У древесных однодольных широко распространены одноосные архитектурные модели (Holttum) и модели с эквивалентными осями (Tomlinson, Chamberlain, реже Leeuwenberg, Schoute) [10]. Модели с гетерогенными скелетными осями встречаются крайне редко (например, у представителей семейства Pandanaceae). При этом число соподчиненных порядков осей незначительно. У однолетних однодольных преобладает изомодульная конструкция, которая также не связана с дифференциацией осей [5].
В крупных таксонах мохообразных структуры с более высокой степенью специализации и дифференциации распространения не так однозначно, как в рассмотренных выше классах цветковых растений. У мхов (отдел Bryophyta) различные варианты дифференциации и специализации структур реализованы более полно, чем у печеночников (отдел Marchantiophyta). Тело всех представителей отдела Bryophyta имеет побеговое строение, характерны многоклеточные ризоиды (таблица) [9]. Только у мхов представлены проводящие ткани с гидроидами и лептоидами, а также механические ткани из стереид. В некоторых группах отмечены варианты строения гаметофита с проводящими тканями в стебле и листьях, образующими единую проводящую систему. Коробочка спорогона мхов нередко имеет достаточно сложное анатомическое строение. В эпидерме могут быть представлены устьичные аппараты. Как правило, в коробочке развивается колонка. Для зеленых мхов (класс Bryopsida) характерно также наличие сложно устроенного перистома. Протонемы у них в большинстве случаев хорошо развиты. Наиболее широкое распространение получили нитчатые протонемы [9].
У печеночников (отдел Marchantiophyta) есть формы с талломным и побеговым строением. В некоторых группах встречаются только талломные формы. Отсутствуют представители с проводящими и механическими тканями. Ризоиды всегда одноклеточные. Спорогоны имеют более простое строение. Ткани в стенках коробочки не дифференцированы, колонка отсутствует [9]. В пределах коробочки нет структур, обеспечивающих порционное распространение спор.
В небольшом по сравнению с мхами и печеночниками отделе Anthocerotophyta анатомическое и морфологическое строение гаметофита достаточно простое. Встречаются только талломные формы. Ткани дифференцированы слабо, нет специализированной ассимиляционной ткани, ризоиды одноклеточные. Однако стенка коробочки спорогона дифференцирована на ткани, в эпидерме встречаются устьичные аппараты, есть колонка. Таким образом, структуры с признаками дифференциации и специализации имеют различное значение в формировании архетипов этих отделов. В пределах классов, представляющих эти отделы, также можно выделить группы с разным характером распространения структур более простого и сложного строения с различным относительным уровнем дифференциации и специализации (таблица).
В двух наиболее крупных по объему таксонах мохообразных (классы Bryopsida Jungermanniopsida) традиционно выделяют группы, характеризующиеся разными типами расположения архегониев. Архегонии, а позднее и спорогон, могут располагаться на верхушке главной, а также равноценной ей оси (класс Bryopsida, группа Acrocarpae) или на верхушке специализированных веточек второго, третьего и более высоких порядков ветвления (класс Bryopsida, группа Pleurocarpae). Отмеченные типы расположения сопряжены с дифференциацией осей в системе побегов. У юнгерманниевых печеночников топография архегониев и их морфогенез в одном случае не препятствуют дальнейшему росту оси (класс Jungermanniopsida группа Anacrogynae), а в другом случае делают этот рост невозможным (класс Jungermanniopsida, группа Acrogynae). Эти особенности также сопряжены с разным уровнем дифференциации осей тела (таблица). В этой связи анализ встречаемости структур с признаками дифференциации и специализации в отмеченных группах представляет специальный интерес.
Для бокоплодных мхов (Bryopsida, группа Pleurocarpae) характерна четкая дифференциация побегов по длительности роста и выполняемым функциям (таблица). Листья на осях разных порядков также могут отличаться размерами и строением. Нередко формируются сложноразветвленные системы осей. Архитектурные модели бокоплодных мхов могут быть сходны с некоторыми моделями древесных растений [8, 11]. Перихециальные листья бокоплодных мхов нередко специализированы в большей степени, чем у верхоплодных. Дифференциация и специализация побегов бокоплодных мхов, как правило, сопряжена с формированием специфических латеральных структур (парафиллии, псевдопарафиллии), способностью образования ризоидов в основании и на дорзальной стороне листа (таблица) [1]. Однако отмеченные выше морфологические особенности у многих бокоплодных мхов сочетаются с относительно невысоким уровнем анатомической дифференциации.
Для верхоплодных мхов (Bryopsida, группа Acrocarpae) характерно отсутствие дифференциации побегов и листьев. Однако в этой группе сформировалось более значительное структурное разнообразие на анатомическом уровне. Представлены различные варианты дифференциации клеток осевых элементов и листьев. Почти у всех бокоплодных мхов клетки листа проземхимные, жилка обычно развита слабо.
У верхоплодных мхов клетки листа, как правило, изодиаметрические, жилка развита хорошо [1]. Только у верхоплодных мхов встречаются варианты строения стебля и листьев с четко дифференцированными проводящими тканями. У представителей семейства Polytrichaceae в пределах листа имеются ассимиляционные пластины, формирующие ассимиляционную ткань, массивное основание с гидроидами, лептоидами и стереидами и структура, похожая на влагалище. Листья с ассимиляционными пластинами встречаются у представителей семейства Pottiaceae. Достаточно сложное морфологическое строение имеют также листья в семействе Fissidentaceae.
Некоторые аналогичные тенденции можно отметить у печеночников из класса Jungermanniopsida. «Акрогинность» также сопряжена с функциональной дифференциацией осей. Акрогинные юнгерманниевые печеночники имеют более высокий уровень морфологической дифференциации тела в целом. Для них характерно побеговое строение, четко выраженная дифференциация осей, которая сочетается со значительным разнообразием типов ветвления, листьев и защитных структур, образующихся вокруг группы гаметангиев и формирующегося спорогона (таблица) [9]. В результате различных структурных преобразований осевой части побега могут формироваться перигинии, марсупии, целокаулы. Специализация перихециальных листьев привела к появлению периантиев различного строения. В пределах листа нередко выражена специализация одной из лопастей. Она преобразовывалась в различные кармановидные или кувшинчатоподобные структуры (таблица). Встречаются листья с перистыми сегментами.
«Анакрогинность» предполагает, как правило, отсутствие функциональной дифференциации осей и сопряжена с более низким уровнем морфологической специализации. Тело имеет талломное строение. Защитные структуры менее разнообразны.
Разный уровень специализации и дифференциации структур гаметофита характерен для анакрогинных юнгерманниевых печеночников и представителей класса Marchantiopsida. Маршанциевые имеют талломное строение. У них дифференцированы подставки, которые могут иметь различную морфологическую природу. В ряде случаев подставки являются метаморфизированной лопастью таллома и имеют сложное анатомическое строение. Слоевище маршанциевых дифференцировано на основную, ассимиляционную и покровную ткани (таблица) [9]. Ассимиляционная ткань может быть сложно структурирована. В этом случае она представляет совокупность ассимиляционных камер, которые ограничены эпидермой, основной тканью и стенками, состоящими из клеток, лишенных хлоропластов. Иногда ассимиляционная ткань имеет ячеистое строение. Ризоиды у Marchantiopsida двух типов – простые и язычковые. Для маршанциевых характерно разнообразие защитных структур. Элатеры могут группироваться в пределах элатофора. По сравнению с маршанциевыми таллом анакрогинных юнгерманниевых печеночников имеет более простое морфологическое и анатомическое строение. Отсутствуют подставки, не дифференцирована ассимиляционная ткань. Образуются только простые ризоиды (таблица). Только у некоторых представителей анакрогинных Jungermanniopsida выражена дифференциация осей таллома (например, у видов рода Aneura Dumort).
Заключение
Дифференциация и специализация были основными модусами структурной эволюции высших растений. Эти модусы реализовывались на морфологическом и анатомическом уровнях. В крупных таксонах выявляются некоторые общие тенденции преобразования частных морфогенетических программ и всей программы онтоморфогенеза. Роль модусов дифференциации и специализации в структурной эволюции разных групп высших растений различна.
Двудольные покрытосеменные растения (Magnoliopsida) отличаются от однодольных (Liliopsida) по полноте реализации модусов дифференциации и специализации на уровне архитектуры тела, отдельных структур, анатомии осевых органов. В отделах мохообразных распространение структур с признаками более высокой специализации и дифференциации не однозначно. По ряду признаков отдел Bryophyta характеризуется более высоким относительным уровнем специализации, чем отдел Marchantiophyta. Однако у верхоплодных мхов при более простой архитектуре нередко наблюдается более сложное анатомическое строение. У маршанциевых таллом морфологически и анатомически дифференцирован в большей степени, чем у анакрогинных юнгерманниевых печеночников. В пределах классов мохообразных выявляется некоторая корреляция встречаемости дифференцированных и специализированных структур с типом расположения архегониев.
Анализ характера реализации модусов структурной дифференциации и специализации дает возможность конструировать архетипы систематических групп, оценивать эволюционную специфику таксонов. При выявлении архетипов таксонов высших растений, объединяющих представителей с модульным типом организации, актуален поиск общих особенностей алгоритмов развития, которые проявляются на разных уровнях структурной иерархии.
Библиографическая ссылка
Нотов А.А. ДИФФЕРЕНЦИАЦИЯ И СПЕЦИАЛИЗАЦИЯ СТРУКТУР В ЭВОЛЮЦИИ НЕКОТОРЫХ ГРУПП ВЫСШИХ РАСТЕНИЙ // Успехи современного естествознания. 2015. № 3. С. 146-150;URL: https://natural-sciences.ru/en/article/view?id=34753 (дата обращения: 11.07.2026).