


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
THE MAPPING STUDY OF THE PLANT COMMUNITIES OF THE ENVIRONMENTAL CONTACT SITES

Составной частью геоботанических исследований является картографическое изучение растительности, выступающее и как метод и как результат одновременно. Картографическое изучение растительности предопределяет выбор подхода в классификации, что, собственно, и является сутью проводимых исследований. Достаточно детальную характеристику подходов в классификации растительности (а без этого невозможно какое-либо картирование сообществ) приводит в своей статье Л.П. Рысин [10] на основе анализа широкого спектра мнений исследователей. От цели проводимых исследований зависит выбор подхода в классификации растительных сообществ. В этой связи уместно привести высказывание Р.В. Камелина по вопросу использования тех или иных классификаций растительности «… будут ли они сколько-нибудь полно востребованы в практике хозяйственной деятельности» [5, с. 565]. Выбор подхода в классификации растительности и как результат исследований – составление карты всегда носит индивидуальный характер и отражает отношение исследователя к той или иной из существующих школ. Мы в наших исследованиях придерживаемся доминантного подхода в классификации растительности В.Н. Сукачева [15], развитого в дальнейшем Е.М. Лавренко [6] и В.Б. Сочавой [14]. При доминантном подходе растения – доминанты являются основой сообществ, определяют их состав и структуру, динамику и деструктивные процессы, происходящие при определенных внешних воздействиях, и отражают условия формирования конкретного сообщества. Направленность эндоэкогенеза и экзоэкогенеза в растительности всегда отражается в составе растений – доминантов (в ряде случаев эдификаторов) ярусов любого сообщества. При использовании доминантного подхода в классификации, помимо того, что максимально учитываются условия экотопов, выделенные таксоны «…легко картографируются и могут быть применены для разработки хозяйственных мероприятий разного целевого назначения» [10, с. 448]. А это и является целью исследований растительности любой территории, где собственно карты растительности разного масштаба лежат в основе геоботанического районирования и являются инструментом в определении типов и форм хозяйственного использования растительного покрова.
В Байкальской Сибири представлен широкий спектр сообществ, отражающих специфику структурно-динамической организации растительного покрова обширных территорий. Такие сообщества могут выступать региональной моделью индикации существующих процессов и прошедших изменений на протяжении последних столетий. Эти изменения могут характеризовать тенденции возможной спонтанной динамики и антропогенных деструкций среды при определенных условиях. Растительные сообщества современного облика, сформировавшиеся на протяжении позднего голоцена, и будут выступать точкой отсчета характера изменений во времени и пространстве. Здесь всегда будет актуальным вопрос: какова природа зональности, поясности, экстразональности или генезис среды, определяющей зональность и экстразональность в организации растительности. На современном этапе это имеет глобальный характер как актуальная проблема современной биогеографии и экологии. Важность картографического изучения растительности контрастных природных условий заключается в том, что растительные сообщества формируются в сложных (контрастных) природных условиях, обусловленных экстразональными эффектами в структуре растительности таежной зоны, в данном случае Западного Прибайкалья.
Целью наших исследований стало определение (методом картографирования) основных особенностей структурно-динамической организации сообществ, формирующихся в условиях взаиморазвития экстразональной степи в границах зональной тайги с определением современных тенденций их генезиса в условиях меняющейся климатической обстановки и вектора влияний антропогенных факторов в регионе. При этом решались следующие задачи:
1) выявить структуру сообществ, формирующихся в условиях взаиморазвития лесов и экстразональной степи,
2) оценить динамические тенденции сообществ контакта лесов и экстразональной степи,
3) определить основные факторы среды, обусловившие формирование сообществ, образованных таежными и степными видами растений,
4) установить типологический состав сообществ, формирующихся в условиях взаиморазвития лесов и экстразональной степи,
5) на основе составленных карт растительности провести прогнозные построения развития сообществ контакта сред на фоне динамики климата и характера влияний антропогенных факторов в регионе.
При классификации степей в целях районирования Е.М. Лавренко [7] под зональной растительностью понимал однородные по составу растительные сообщества плакоров. Им было проведено районирование степной зоны на зонально-провинциальные подразделения, где зона степей рассматривается как самостоятельное природное явление с характеристикой региональных неоднородностей структуры растительности. Занимаясь проблемой зонирования природной среды СССР, Л.С. Берг [4] под природными зонами понимал области преобладающего развития одних и тех же ландшафтов. Им были выделены тундровая, лесная, лесостепная, полупустынная, пустынная и субтропическая зоны. И здесь мы видим вполне определенные характеристики, определяющие ту или иную зону. По мнению В.Б. Сочавы [13, 14], географическая зональность – это главнейшее свойство географической оболочки, обусловливающее дифференциацию среды определенного качества. В таежной зоне Сибири встречаются участки степей в виде «степных островов» различной площади, часто по межгорным впадинам, по долинам рек, крутым склонам, иногда достигая границы тундры. Они не создают природных зон в пространственном отношении, как, например, лесостепные и подтаежные [17]. С севера на юг соотношение между тайгой и степью изменяется в пользу последней. Встречаемость остепненных участков среди тайги уменьшается с востока на запад.
В отличие от зональной лесостепи и подзональной (плакорной) подтайги, а также лесостепного пояса и пояса низкогорной подтайги в горах, «островные степи» являются в некотором роде сукцессией зонального типа растительности или горного пояса со своей спецификой структуры и генезиса сообществ. В горных условиях взаимоотношение тайги и степей характеризуется формированием горностепного пояса [15]. Для горных стран характерна пространственная и вертикальная стратиграфия структуры растительности в условиях зональности и высотной поясности, которая отражает конкретные физико-географические условия среды. Это не вызывает сомнений. Сомнение вызывает то, что одним и тем же термином характеризуются разные природные явления (или среды). При оценке развития и формирования растительных сообществ необходим всесторонний учет специфики территории с целью избежания неопределенности в понимании того, какое явление подразумевается в каждом конкретном случае. Необходимо конкретизировать смысловые нагрузки терминов, что есть лесостепь, подтайга в условиях зональности, высотной поясности, и, главное, в условиях экстразональности. Эти природные явления отражают совершенно конкретные физико-географические условия территории. Вероятно, следует как-то разграничивать информационную сущность терминов, характеризующих конкретное природное явление.
В настоящее время накоплен огромный массив публикаций, характеризующих растительные сообщества как «экотоны». Однако экотоны в структуре растительности выделяют и в условиях зональности и в условиях экстразональности растительности и как опушка леса. То есть происходит инверсия данного термина, используемого и для характеристики межзональных (или межпоясных) и внутризональных (или внутрипоясных) растительных сообществ разных по физико-географическим условиям территорий. Впервые термин «экотон» был использован в работах английских и американских исследователей [18, 19] как характеристика перехода между двумя растительными ассоциациями. Ими указывается, что «экотон» – переход между двумя сообществами растений с повышенным видовым богатством в зонах перехода смежных участков растительности. Большую роль экотонов как буферных сообществ в организации растительности учитывал в своих исследованиях В.Б. Сочава [13]. Он утверждал, что там, где есть экотон, создается впечатление расплывчатости границ между сообществами, а показ на картах экотона несколько устраняет трудности проведения границ, однако одновременно это вызывает ряд новых вопросов, касающихся отражения структуры и генезиса растительных сообществ. Применение термина «экотон» для разных физико-географических условий (зональных, высотно-поясных, экстразональных) вряд ли является оптимальным решением вопроса классификации растительных сообществ контакта сред. Характеризовать разнокачественные объекты разных условий одним и тем же термином – значит вносить путаницу в установлении того, какое явление рассматривается. «Опушечный», «межзональный», «зональный» или «экстразональный» экотоны – это определенно разнокачественные явления, отражающие разные среды. Использование того или иного термина всегда связано с глубиной знания конкретных объектов, их классификационной принадлежностью к конкретному природному явлению. Решить проблему типизации и классификации растительных сообществ контакта сред, определить их информационное содержание возможно методом крупномасштабного картографирования растительных сообществ на регионально-топологическом уровне организации растительности.
Материалы и методы исследования
Ключевым участком наших исследований выступила территория формирования зональных лесов и экстразональной («островной») степи центральной части западного побережья оз. Байкал (бассейн р. Таловки и побережье Малого Моря). Они оределены так, чтобы выявить структурно-пространственную организацию растительности весьма контрастного по физико-географическим условиям всего западного побережья оз. Байкал.
Предметом исследований является особенности взаимоотношений и взаиморазвитие светлохвойной зональной тайги и экстразональной степи, обусловивших формирование растительных сообществ, образованных видами растений разной ареалогии и экологии. В результате геоботанической съемки разных лет проведено более 900 геоботанических описаний разных лет наблюдений в комплексе со сборами гербарного материала. Составленые картосхема и карта растительности крупного масштаба, позволили выявить особенности пространственной стратиграфии сообществ контакта зональной тайги и экстразональной степи. В целях проведения полномасштабных исследований растительности сложных (контрастных) физико-географических условий территории был необходим комплекс методов, которые позволили выявить детальную структурно-динамическую организацию растительности контрастных природных условий. При проведении геоботанической съемки учитывались рекомендации, изложенные в научной литературе [8, 9]. Крупномасштабное картографирование проводилось с использованием материалов аэрокосмофотосъемки [1]. Закладка модельных площадок проведена в целях мониторинга внутрисезонной и межсезонной динамики растительных сообществ в целях отражения динамической их составляющей при создании инвентаризационных картосхем и собственно карт растительности. При типизации сообществ были использованы конкретные принципы классификации растительности, озвученные в научной литературе [6, 14, 15].
Проведение работ осуществлялось поэтапно: 1-й этап – типизация и классификация растительных сообществ, морфологически, экологически и динамически отражающих зональные (или экстразональные), регионально-типологические особенности современной структуры растительности района исследований; 2 – создание крупномасштабных картографических моделей (картосхем) пространственной организации сообществ ключевого участка с учетом строения рельефа, эдафических условий и пространственного положения сообществ в системе гор Прибайкалья; 3 – типизация и классификация растительных группировок, морфологически, экологически и динамически отличных от ассоциаций, отражающих определенный тип растительности, с созданием картографических моделей (картосхем); 4 – создание системы модельных участков (каркаса), репрезентативно отражающих все разнообразие растительных сообществ территории исследований; 5 – выявление характера связей между составляющими ценоз компонентами, определяющими растительное сообщество как систему; 6 – установление параметров, определяющих конкретное состояние сообщества, которые могут быть использованы для экстраполяции в оценке подобных (близких к моделям) сообществ других районов; 7 – составление собственно карты растительности. При этом всесторонне учитывался антропогенный фактор, влияющий на структурно-динамическую организацию конретного сообщества.
Геоботаническая съемка на базе полевого дешифрирования крупномасштабных аэрофотоснимков разных лет выявила пространственно-структурные связи растительных сообществ, особенности и направленность динамических процессов в растительности, а также фитоиндикационные свойства сообществ (видов), обусловившие мозаичность и синузиальность растительного покрова ключевых участков района исследований. Этому предшествовал этап камеральных работ с составлением картосхем на основе разностей в пространственной структуре изображений растительности на аэрофотоснимках. При картосоставлении типизация и классификация сообществ осуществлялась в следующей последовательности: получение набора описаний участков растительности (лесных и степных ценозов отдельно) с отметкой на снимке точек описания – группировка описаний по сходству состава доминантов ярусов и горизональной структуре – группировка описаний по сходству местообитаний сообществ – классификация с определением ассоциаций. В результате была составлена крупномасштабная базовая инвентаризационная картосхема растительных сообществ ключевого участка западного побережья оз. Байкал.
Результаты исследований и их обсуждение
Крупномасштабное картографическое изучение растительного покрова переходных (межзональных, межвысотно-поясных, внутризональных, азональных) территорий позволило выявить структурно-динамические особенности растительности на регионально-топологическом уровне ее организации. При этом понятие зональности или экстразональности в растительности конкретизировались факторальными рядами, обусловившие структуру растительности конкретной территории. Выявленные внутрисистемные связи в процессе формирования сообщества переходного типа (контакта сред) позволили определить современные динамические тенденции, провести реконструкцию растительности прошлого, выстраивать прогнозы потенциальных изменений в сообществах. Картографический анализ особенностей структуры сообществ контрастных условий позволил выявить механизмы формирования таких сообществ, как мест формирования видового и ценотического разнообразия трансграничных территорий. Картографическое отображение сообществ, состоящих из разных экобиоморф и ареалогического состава видов растений, позволило выявить процессы, отражающие формирование пространственно и генетически особых систем и получить модели в целях мониторинга формирования среды регионально-топологического уровня ее организации.
Район исследований входит в состав административного района Иркутской области – Ольхонского, охватывающего большую часть западного побережья оз. Байкал. Территория характеризуется высокой контрастностью природных условий со значительной внутригодичной и разногодичной динамикой составляющих среду компонентов, главным образом осадков и температуры, и входит в состав особо охраняемой природной территории – Прибайкальского национального парка. Актуальность в этой связи приобретает изучение и выявление детальной структурно-динамической организации растительности, как наиболее динамичного компонента природных систем, аккумулирующего и отражающего практически все изменения в вертикальной и горизонтальной стратиграфии природной среды, ее генезиса.
Растительность района исследований – центральной части западного побережья оз. Байкал (Приольхонье) отличается сложной пространственной организацией сообществ. Здесь развиты светлохвойные таежные леса с доминированием сосны обыкновенной в южной части Приольхонья и лиственницы сибирской побережий Малого Моря оз. Байкал. В пределах этих районов в комплексе с лесами развиты степные сообщества. Здесь среди степей развиты разновозрастные (от 2 до 45 лет) разнотравные остепненные лиственничники практически по всей территории.
Современное состояние растительности определяется общим характером ландшафтно-растительных условий. Специфика растительного покрова центральной части западного побережья отражает определенную связь с историей развития растительности всего Байкальского региона. Основные региональные структурно-типологические особенности и структурно-динамические признаки современного растительного покрова Байкальского региона в целом и территории западного побережья оз. Байкал отражены на мелкомасштабной (м-б 1: 1500 000) геоботанической карте [3]. Она показывает, что для растительного покрова района исследований характерен широкий набор флористически, ценотически и динамически конкретных сообществ от горно-таежных лесов до степных сообществ.
Инвентаризационная крупномасштабная картосхема растительных сообществ контакта зональной тайги и экстразональной степи ключевого участка западного побережья оз. Байкал
Составленная инвентаризационная картосхема (рис. 1) растительных сообществ контакта зональной тайги и экстразональной степи выступила основой для определения ключых участков в целях выявления наиболее типичных сообществ контакта тайги и экстразональных степей для центральной части западного побережья оз. Байкал (рис. 2).
Составленная инвентаризационная картосхема на бассейн р. Таловки (рис. 1) послужила основой для характеристики особенностей формирования сообществ контакта тайги и экстразональной степи района исследований. Картосхема сообществ ключевого участка позволила провести дальнейшие классификационные построения. Сообщества, сходные по составу экобиоморф-доминантов ярусов и типам местообитаний объединялись в ассоциации в целях составления собственно геоботанической карты растительности.
Здесь необходимо отметить, что во флоре сообществ контакта зональной тайги и экстразональной степи отмечено 674 вида высших сосудистых растений, относящихся к 68 семействам 287 родам и мхов – 32 вида. Соотношение основных групп высших сосудистых растений в целом характерно для голарктических флор. На долю 11 ведущих семейств (Poaceae, Asteraceae, Cyperaceae, Rosaceae, Fabaceae, Ranunculaceae, Caryophyllaceae, Brassicaceae, Apiaceae, Scrophulariaceae и Lamiaceae) приходится 431 вид растений, что составляет 67,8 % от общего числа, а на долю остальных 57 семейств приходится 243 видов растений, что составляет 32,2 % от общего состава. Такой набор ведущих семейств характерен для бореальных флор и схож с семейственным спектром для флоры всей Восточной Сибири. Для всей голарктической области очень показательно ведущее положение семейств Asteraceae, Poaceae. Для бореальных флор весьма характерна высокая роль Cyperaceae, значительная – Ranunculaceae. Континентальные черты флор проявляются в значительной роли семейств Brassicaceae, Rosaceae, Fabaceae. Относительное обилие видов Caryophyllaceae, Scrophulariaceae возрастает к северу и достигает максимума в арктической полосе Сибири. Семейственный спектр воплощает наиболее общие особенности флоры в связи с ее зональным положением (зона тайги).
В основу выделенных типов геоэлемента (типов ареалов) положены принципы, изложенные в работах многих исследователей. Всего выделено 14 типов геоэлемента. В составе флоры соообществ контакта тайги и степей западного побережья оз. Байкал преобладают представители бореальной, южносибирской, евразиатской флор, тогда как количество видов центральноазиатской и общеазиатской флор незначительно.
На принципах деления видового состава по признакам приуроченности к той или иной поясно-зональной группе проведен анализ флоры сообществ контакта тайги и экстразональной степи западного побережья оз. Байкал. Показано, что из всего видового состава к лесостепной поясно-зональной группе относятся 89 видов (относящихся к 22 семействам), что составляет 13,17 % от всего видового состава (674 вида). К собственно степной группе относится 63 вида (относящихся к 15 семействам), что составляет 10,04 % от общего видового состава сообществ контакта тайги и экстразональной степи западного побережья оз. Байкал. Присутствие той или иной группы видов, характерных для конкретного высотного пояса (или природной зоны) всегда отражает сущность явления, его среду. Отсюда следует, что в районе исследований нет ярко выраженной поясной структуры и характеризовать сообщества контакта тайги и экстразональной степи как горно-лесостепной или степной пояса вряд ли является правомерным.
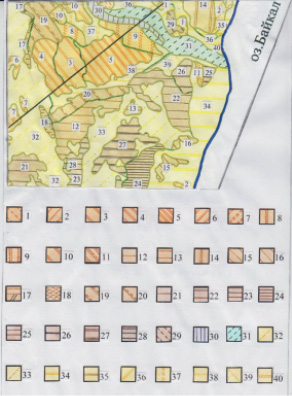
Рис. 1. Фрагмент картосхемы сообществ контакта зональной тайги и экстразональной степи. Ключевой участок – бассейн р. Таловки, южная часть Приольхонья (м-б 1: 25 000)
ЛЕГЕНДА
К КАРТОСХЕМЕ РАСТИТЕЛЬНЫХ СООБЩЕСТВ КЛЮЧЕВОГО УЧАСТКА БАССЕЙНА Р. ТАЛОВКИ
М I: 25 000
Сосновые (Pinus sylvestris L.) леса
1. Сосновые с подростом сосны разнотравно (Phlomis tuberosa L., Galium verum L.) – злаковые (Festuca lenensis Drob.) остепненные леса на эродированных склонах.
2. Сосновые с подростом сосны петрофитные редкотравные (Youngia tenuifolia Willd., Orоstschys spinosa (L.) С.A. Mey.) остепненные леса на эродированных склонах, обращенных к Байкалу (крутые обрывистые склоны).
3. Сосновые с подростом сосны петрофитные остепненные (Astragalus versicolor Pall., Festuca lenensis) леса на слабоэродированных склонах.
4. Cосновые c подростом сосны петрофитные остепненные (Poa botryoides (Trin. ex Griseb.) Roshev., Orostachys spinosa) леса на слабоэродированных склонах.
5. Сосновые с подростом сосны петрофитные остепненные (Heteropappus altaicus (Willd.) Novopokr., Artemisia gmelinii Web. ex. Stechm.) леса на слабоэродированных склонах.
6. Сосновые с лиственницей с подростом сосны и лиственницы остепненные (Phlomis tuberosa, Leontopodium ochroleucum subsp. campestre (Ledeb.) V. Khan.) леса на шлейфах эродированных склонов разлома.
7. Сосновые с лиственницей, березой с подростом сосны остепненные (Artemisia frigida Willd., Phlomis tuberosa) леса на выположенных склонах разлома.
8. Сосновые с лиственницей с подростом сосны, лиственницы, березы спирейные (Spiraea media Fr. Schmidt) осоковые (Carex macroura Meinsh.) ксеромезофитные леса на эродированных склонах.
9. Сосновые с лиственницей с подростом сосны, лиственницы осоковые (Carex macroura) спирейные (Spiraea media) ксеромезофитные леса на эродированных склонах.
10. Сосновые с подростом сосны петрофитные спирейные разнотравные (Phlomis tuberosa, Galium verum) злаковые мезофитные леса на эродированных склонах.
11. Сосновые с подростом сосны петрофитные (Astragalus versicolor, Chamaerhodos altaica (Laxm.) Bunge) со спиреей (Spiraea media) мезофитные леса на эродированных склонах.
Лиственнично (L. sibirica Ledeb.) – сосновые (P. sylvestris) леса
12. Лиственнично-сосновые с подростом лиственницы и сосны разнотравные (Aster alpinus L., Galium verum) остепненные леса широких водоразделов.
13. Лиственнично-сосновые с подростом лиственницы и сосны разнотравные остепненные (Artemisia frigida, Festuca lenensis) леса широких водоразделов.
14. Лиственнично-сосновые с подростом сосны, лиственницы петрофитные остепненные (Koeleria cristata (L.) Pers., Agropyron cristatum (L.) Beauv.) леса на эродированных склонах.
15. Лиственнично-сосновые с подростом сосны, лиственницы петрофитные редкотравные (Youngia tenuifolia, Orоstachys spinosa) остепненные леса по эродированным склонам, обращенным к Байкалу (крутые обрывистые склоны).
16. Лиственнично-сосновые с подростом сосны, лиственницы петрофитные (Artemisia gmelinii, Chamaerhodos altaica) остепненные (Festuca lenensis) леса склонов береговой линии оз. Байкал.
17. Лиственнично-сосновые с подростом сосны, лиственницы разнотравные (Potentilla bifurca L., Vicia cracca L.) мезофитные леса на седловинах.
18. Лиственнично-сосновые с подростом лиственницы и сосны с березой разнотравные (Potentilla tanacetifolia Willd ex Schlecht., Vicia cracca, Pulsatilla flavescens (Zucc.) Juz.) мезофитные леса седловин.
19. Лиственнично-сосновые с подростом сосны, лиственницы осоковые (Carex macroura) мезофитные леса на слабоэродированных склонах.
20. Лиственнично-сосновые с подростом сосны, лиственницы осоковые (Carex macroura) мезофитные леса на слабоэродированных склонах.
Сосново (Pinus sylvestris) – лиственничные (Larix sibirica) леса
21. Сосново-лиственничные с подростом лиственницы, сосны кизильниковые (Cotoneaster melanocarpus Fisch. et Blytt) осоковые (Carex macroura) мезофитные леса на широких водоразделах.
22. Сосново-лиственничные с осиной, сосной с подростом лиственницы разнотравно (Galium verum, Potentilla tanacetifolia, Pulsatilla flavescens)-осоковые (Carex macroura) с участием кизильника черноплодного мезофитные леса широких водоразделов.
23. Сосново-лиственничные с сосной, лиственницей с подростом лиственницы и сосны разнотравно (Lathyrus humilis (Serg.) Spreng., Pulsatilla flavescens, Iris ruthenica Ker-Gawler) осоковые (Carex macroura) мезофитные леса широких водоразделов.
24. Сосново-лиственничные с подростом лиственницы кизильниковые осоковые (Carex macroura) мезофитные леса широких водоразделов.
25. Сосново-лиственничные с подростом лиственницы, сосны, осины петрофитные злаковые (Poa botryoides) ксеромезофитные леса на водоразделах.
26. Сосново-лиственничные с подростом лиственницы, сосны петрофитные разнотравные (Galium verum, G.boreale L., Androsace incana Lam.) мезофитные леса на водоразделах.
27. Сосново-лиственничные с подростом лиственницы и сосны разнотравные (Phlomis tuberosa, Iris ruthenica) с осокой (Carex macroura) и участием кизильника черноплодного мезофитные леса на водоразделах.
28. Сосново-лиственничные с подростом сосны, лиственницы петрофитные осоковые (Carex macroura) разнотравные c синузиями мхов (Rhytidium rugosum (Hedw.) Kindb., Abietinella abietina (Turn.) Fleisch. мезофитные леса на водоразделах.
29. Сосново-лиственничные с подростом лиственницы, сосны спирейные (Spiraea media) осоково (Carex mocroura)-зеленомошные (Dicranum polysetum Sw., Pleurozium schreberi (Brid.) Mitt.,Hyloconimum splendens (Hedw.) B.S.G. мезофитные леса на эродированных склонах.
Елово (Picea obovata Ledeb.) – лиственничные (Larix sibirica) с березой
(Betula pendula Roth) и осиной (Populus tremula L.) леса надпойменных террас долины р. Таловки
30. Березово-осиновые с лиственницей и елью с подростом ели, лиственницы с ивами (Saliх myrtilloides L., Salix rorida Laksch.) широкотравно (Aconitum baikalense Turcz. ex Rapaics) – осоковые (Carex bicolor All., C. enervis C.A. Mey.) леса низких надпойменных террас.
Луговые сообщества
31. Злаковые (Alopecurus pratensis L., Agrostis gigantea Roth) осоковые, (C. coriophora Fisch. et Mey. ex Kunth) и пушицевые (Eriophorum russеolum Fries) заболоченные луга низких надпойменных террас.
Сообщества экстразональных степей
32. Осоковые (Carex duriuscula C.A. Mey.) с разнотравьем (Potentilla bifurca, Phlomis tubirosа) сообщества седловин широких водоразделов.
33. Разнотравные (Heteropappus altaicus, Veronica incana L.) с тонконогом (Koeleria cristata) сообщества на седловинах широких водоразделов.
34. Разнотравные (Youngia tenuifolia, Orostachys spinosa) тимьяновые (Thymus baikalensis Serg.) петрофитные сообщества на эродированных склонах, обращенных к Байкалу (крутые обрывистые склоны).
35. Злаковые (Agropyron cristatum, Poa botryoides) сообщества на эродированных склонах прибрежья оз. Байкал.
36. Тимьяновые (Thymus baikalensis) c овсянницей (Festuca lenensis) сообщества на эродированных склонах.
37. Злаковые (Festuca lenensis, Agropyron cristatum) петрофитные мезоксерофитные сообщества на эродированных склонах.
38. Разнотравно (Phlomis tuberosa, Veronica incana) – злаковые (Phleum phleoides (L.) Karsten, Festuca lenensis) cообщества по шлейфам эродированных склонов.
39. Тимьяновые (Thymus baikalensis) с осокой (Carex duriuscula) разнотравные (Leontopodium ochroleucum subsp. campestre) мезоксерофитные сообщества по шлейфам эродированных сконов.
40. Разнотравно (Veronica incana, Echinops latifolia Tausch.) – тимьяновые (Thymus baikalensis) сообщества по шлейфам эродированных склонов.
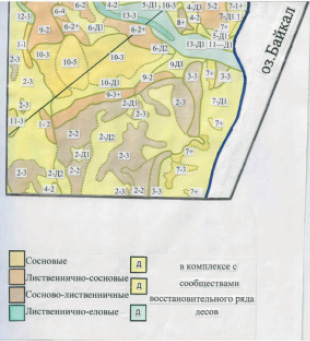
Рис. 2. Фрагмент карты таежно-степных сообществ (эпиассоциаций) ключевого участка «Бассейн р. Таловки», южная часть Приольхонья (м-б 1: 25 000)
ЛЕГЕНДА
К КАРТЕ ТАЕЖНО-СТЕПНЫХ СООБЩЕСТВ (ЭПИАССОЦИАЦИЙ) КЛЮЧЕВОГО УЧАСТКА БАССЕЙНА РЕКИ ТАЛОВКИ
М 1: 25 000
Южно-Сибирские формации
Эпиформации светлохвойных (Рinus sylvestris L., Larix sibirica Ledeb.) горнотаежных лесов
Эпиассоциации таежно-степных сообществ
1. Сосновые (Pinus sylvestris) с лиственницей (Larix sibirica), лиственнично-сосновые рододендроновые (Rhododendron dauricum) с душекией (Duschekia fruticosa (Rupr.) Pouzar брусничные (Vaccinium vitis-idaea L.) мезофитные леса на седловинах водоразделов.
1–1. Осиново (Populus tremula L.)-березовые (Betula pendula Roth) леса.
1–2. Лиственнично-сосновые с березой, осиной с подростом леса.
2. Сосново (Pinus sylvestris) – лиственничные (Larix sibirica) разнотравные (Iris ruthenica Ker-Gawler, Scorzonera radiata Fisch., Sanguisorba officinalis L.) остепненные (Bupleurum scorzonerifolium Willd., Heteropappus altaicus (Willd.) Novopokr., Phlomis tuberosa L., Aster alpinus L., Scabiosa comosa Fisch. ex Roemer et Schult., с участием Cotoneaster melanocarpus Fisch. et Blytt, и Carex macroura Meinsh., и Iris ruthenica леса на водоразделах.
2–Д1. травянистые с доминированием гетеропаппуса алтайского, Veronica incana L., Leontopodium ochroleucum subsp. сampestre (Ledeb.) V. Khan. бледножелтого ксерофитные сообщества
2–3. Сосново-лиственничные леса с осиной, березой с подростом, с кизильником черноплодным, осокой большехвостой, ирисом русским.
2–Д2. Травянистые с доминированием Potentilla tanacetifolia Willd. ex Schlecht., володушки козелецелистной, гетеропаппуса алтайского, эдельвейса обыкновенного (Leontopodium leontopodioides (Willd.) Beauv.), Koeleria cristata (L.) Pers., Astragalus versicolor Pall. ксерофитные сообщества.
2–2. Сосновые с лиственницей, осиной, березой без подроста кизильниковые осоковые ксеромезофитные леса.
3. Сосново (Pinus sylvestris)-лиственничные (Larix sibirica) и лиственничные с сосной петрофитные редкотравные (Lupinaster pentaphyllus Moench, Scorzonera radiata, Artemisia gmelinii Web. et Stechm., Aster alpinus, Dianthus versicolor Fisch. ex Link) мезофитные леса на эродированных склонах.
3–3. Сосново-лиственничные с подростом леса.
4. Сосновые (Pinus sylvestris) с лиственницей (Larix sibirica) разнотравные (Phlomis tuberosa, Veronica incana, Heteropappus altaicus остепненные леса на эродированных склонах.
4–5. Березово-осиновый подрост на вырубках.
4–4. Сосновые леса с лиственницей, березой с подростом сосны и лиственницы.
4–3. Сосновые леса с лиственницей, березой с подростом сосны и лиственницы.
4–Д5. Травянистые с доминированием мятлика оттянутого (Poa attenuata Trin.), житняка гребенчатого (Agropyron cristatum (L.) Beauv.) ксерофитные сообщества.
4–2. Сосновые с лиственницей леса, с березой, осиной без подроста.
4–Д3. Травянистые с доминированием полыни холодной (Artemisia frigida Willd.), хамеродоса прямостоячего (Chamaerhodos erecta (L.) Bunge), тимьяна байкальского (Thymus baicalensis Serg.), тонконога гребенчатого ксерофитные сообщества.
5. Сосновые (Pinus sylvestris) с лиственницей (Larix sibirica) и лиственнично-сосновые петрофитные остепненные редкотравные (Astragalus fruticosus Pallas, Artemisia frigida, Galium verum L., Koeleria cristata) леса на эродированных склонах.
5–2. Сосновые с лиственницей, березой леса.
5–Д1. Травянистые с доминированием гетеропаппуса алтайского, вероники седой, Silene jenisseensis Willd. ксерофитные сообщества.
6. Сосновые (Pinus sylvestris) с лиственницей (Larix sibirica) петрофитные редкотравные (Carex macroura) с таволгой (Spiraea media Fr. Schmidt) и участием мхов (Dicranum polysetum Sw., Pleurozium schreberi (Brid.) Mitt., Rhytidium rugosum (Hedw.) Kindb., Dicranum acutifolium (Lindb. et Arn.) C. Jens., Climacium dendroides (Hedw.) Web. et Mohr, Ptilium crista-castrensis (Hedw.) De Not.) мезофитные леса на эродированных склонах.
6–4. Сосновые с лиственницей и подростом леса.
6–2. Сосновые с лиственницей и подростом леса.
6–Д1. Травянистые с доминированием житняка гребенчатого, зопника клубненосного ксерофитные сообщества.
6–2+. Сосновые с лиственницей с подростом сосны, лиственницы и березы леса.
6–Д2. Травянистые с доминированием Carex duriuscula C.A. Mey.
мятлика кистевидного (Poa botryoides (Trin. ex Griseb.) Roshev.) ксерофитные сообщества.
7. Сосновые (Pinus sylvestris) с лиственницей (Larix sibirica) петрофитные редкотравные остепненные (Youngia tenuifolia Willd., Papaver nudicaule L., Artemisia gmelinii, Pulsatilla turczaninovii Krylov et Serg., Lilium pumilum Delile, Allium anisopodium Ledeb., Polygonatum odoratum (Mill.) Druce леса на эродированных склонах береговой линии оз. Байкал.
7–3. Сосновые с лиственницей и подростом леса.
7–1+. Сосновые с подростом сосны и березы леса.
7–Д1. Травянистые с доминированием юнгии тонколистной, Thymus baikalensis, лилии карликовой, полыни Гмелина с участием Ephedra monosperma C.M. Mey и Orostachys spinosa (L.) C.A. Mey, ксерофитные сообщества.
7+. Сосновые с лиственницей с подростом сосны и березы леса.
7–Д1.1. Травянистые с доминированием овсяницы ленской (Festuca lenensis Drob.), хамеродоса прямостоячего, Potentilla bifurca L. ксерофитные сообщества.
8. Сосново (Pinus sylvestris) – лиственничные (Larix sibirica) кустарниковые (Spiraea media) осоково (Carex macroura)-зеленомошные (Climacium dendroides, Dicranum polisetum Sw., Pleurozium scheberii ксеромезофитные леса на эродированных склонах.
8+. Сосново-лиственничные с подростом лиственницы и березы леса.
9. Лиственнично (Larix sibirica)-сосновые (Pinus sylvestris Festuca lenensis Drob.) разнотравные (Lathyrus humilis (Serg.) Spreng., Vicia venosa (Willd. ex Link) Maxim., Geranium pseudosibiricum J. May., Carex macroura) мезофитные леса на слабоэродированных склонах.
9–3+. Лиственнично-сосновые, с подростом сосны, лиственницы и березы леса.
9–Д1. Травянистые с доминированием тонконога гребенчатого, Allium tenuissimum L., осоки твердоватой, лапчатки вильчатой остепненные сообщества.
9–2. Лиственнично-сосновые с березой с подростом леса.
10. Сосновые (Pinus sylvestris) разнотравные (Phlomis tuberosa, Heteropappus altaicus, Potentilla tanacetifollia, Galium verum, Androsace lactiflora Fisch. ex Duby, Orostachys spinosa) петрофильные остепненные леса на слабоэродированных склонах.
10–5. Подрост сосняка на гари.
10–3. Сосновые с подростом леса.
10–Д1. Травянистые с доминированием тонконога гребенчатого.
ксерофитные сообщества.
11. Сосновые (Pinus sylvestris) с лиственницей (Larix sibirica) разнотравные остепненные (Sanguisorba officinalis, Aster alpinus, Galium verum, Phlomis tuberosa, Phleum phleoides (L.) Karsten, Festuca lenensis, Iris humilis Georgi, Veronica incana) леса на шлейфах эродированных склонов.
11–3. Сосновые с лиственницей с подростом леса.
11–Д1. Травянистые с доминированием мятлика оттянутого, тимофеевки степной, истода сибирского (Polygala sibirica L.) с участием ириса низкого.
12. Сосново (Pinus sylvestris)-лиственничные (Larix sibirica) широкотравные (Heracleum dissectum Ledeb., Valeriana alternifolia Ledeb., Veratrum lobelianum Bernh., Geranium wlassowianum Fisch. ex Link, Trollius asiaticus L., Aconitum baikalense Turcz. ex Rapaics) мезофитные по седловинам и U-образным распадкам.
12–3. Сосново-лиственничные с березой с подростом леса.
13. Лиственнично (Larix sibirica)-еловые (Picea obovata Ledeb.) с сосной (Pinus sylvestris) с участием ив (Salix myrtilloides L., S. rorida Laksch.) широкотравные (Cacalia hastata L., Carum carvi L., Trollius asiaticus, Equisetum prаtense Ehrh., Veratrum lobelianum) леса низких надпойменных террас долины р. Таловка.
13-3 березовые с осиной с участием ив разнотравные (Carex coriophora Fisch. et Mey. ex Kunth, C. rynchophysa C.A. Mey., C. bicolor All.) гигромезофитные заболоченные леса.
13-Д1 травянистые сообщества с доминированием (Agrostis gigantea Roth, Alopecurus pratensis L., Equisetum pratense, Hordeum brevisubulatum (Trin.) Link, Carex enervis C.A. Mey., Eriophorum russеolum Fries, Trollius asiaticus, Carum carvi, Veratrum lobelianum).
Дополнительные обозначения к легенде (динамические составляющие эпиассоциацию):
1–1, 1–2, 2–3 – первая цифра означает порядковый номер эпиассоциации, вторая – возрастную группу (5 градаций) лесообразующей породы, где 5 – возраст до 20 лет, 4 – до 40 лет, 3 – до 60 лет, 2 – до 80 лет, 1 – до 100 лет и выше); + – леса с подростом из лесообразующей и мелколиственной породами деревьев; Д1, Д1.1, Д2, Д5 и т.д. – травянистые сообщества демутационно-восстановительного ряда, как составная часть динамики в генезисе лесов в рамках таежно-степных эпиассоциаций.
Используя подходы в классификации видов растений по отношению их к среде, был проведен экотипологический, экоморфологический и эколого-географический анализы флористического состава сообществ контакта тайги и экстразональной степи района исследований. Из общего видового состава сообществ (674 вида) на долю эуксерофитов, криоксерофитов, мезоксерофитов, ксеромезофитов приходится 38 % видов растений. Виды-мезофиты составляют соответственно 52 %. Надо отметить, что часто происходит смена доминирующих позиций в сообществах в разных климатических условиях растений разной экологии. Достаточно широко представлен набор травянистых сообществ с ядром видов-ксерофитов со специфическими условиями местообитаний, резкими сезонными и разногодичными колебаниями климатических фактров, в основном осадков, которые обусловили формирование достаточно устойчивых ценозов в сложившихся условиях географической среды. Большое разнообразие мезофитов (более 50 %) обусловлено влиянием таежного элемента растительного покрова.
По эколого-ценотической приуроченности флоры сообществ контакта тайги и экстразональной степи существенная доля видов растений принадлежит лугово-лесной, луговой и лесной группам растений (48 % от общего состава видов), тогда как к лугово-степной и степной группе относится 42 % видового состава и к лугово-болотной и сорной группам 10 % видов. В составе степных сообществ широко представлены виды растений, характерные для тайги и высокогорья (к примеру, дриада Сумневича (Dryas sumneviczii), проломники – седой, молочноцветковый (Androsace incana, A. lactiflora). Большой процент от общего состава видов принадлежит не степной группе, что вполне закономерно в современных природных условиях и динамических тенденциях формирования растительности региона.
Многолетние исследования растительности методом крупномасштабного геоботанического картографирования позволили выявить некоторые особенности структуры и развития растительных сообществ, формирующихся при взаимовлиянии зональных лесов и экстразональных степных сообществ на пространственно ограниченной территории.
Крупномасштабная карта таежно-степных сообществ (эпиассоциаций) на ключевой участок западного побережья оз. Байкал
Для выявления и отражения всей сложности ценоструктуры таких сообществ был использован структурно-динамический принцип классификации растительности [14], используемый в геоботанической картографии с составлением карт разных масштабов. Основной единицей классификации в данном случае выступает эпиассоциация, как сложная динамическая система, включающая максимально возможные состояния сообществ, образующих растительную ассоциацию для конкретного типа местообитания. Эпиассоциация расматривается как синтаксон, объединяющий разные динамические состояния ассоциации – от серийных до коренных. В эпиассоциацию «…входят все переменные составляющие ассоциации…разного рода серии и ряды развития в направлении материнского ядра ассоциации». Эпиассоциация в нашем понимании оценивается как система, где отражены: возрастная структура древостоев, состав доминантов и содоминантов конкретной возрастной группы, наличие или отсутствие подроста и его состав, присутствие или отсутствие (доминирование) мелколиственных пород, характер естественного возобновления (в зависимости от фактора воздействий – пожар, рубка, выпас, сенокос). Для травянистых (степных) сообществ, входящих в состав эпиассоциации как демутационные или восстановительные ряды, генетически связанные с лесными ценозами, приводится характеристика ярусности, доминантов ярусов и обилия характерных видов, а также условия формирования и генезис сообществ. Для этого была проведена классификация сообществ, которая имеет вид системы последовательной типизации фитоценозов по мере набора описаний сообществ со сходной структурой, динамикой и условиями их местообитаний.
Сообщества, формирующиеся в условиях контакта зональной тайги и экстразональной степи следует называть таежно-степными, которые по структуре и динамике образуют эпиассоциацию для конкретного типа местообитания. Составление крупномасштабной (в м-бе 1: 25 000) карты таежно-степных сообществ (эпиассоциаций) на ключевой участок позволило выявить все разнообразие пространственной структуры и вскрыть особенности вертикальной организации сообществ, формирующихся в процессе взаиморазвития и взаимовлияний светлохвойной тайги и экстразональных степей центральной части западного побережья оз. Байкал. При типизации и классификации сообществ испоьзован доминантный подход [15] с учетом эколого-морфологических признаков [6] и геосистемного анализа структурно-динамических признаков сообществ, объединенных в эпитаксоны [14]. Выявлен большой набор различных по структуре ценозов, образующих сложный комплекс светлохвойных (таежных) и степных сообществ. Основу растительности района исследований образуют эпиассоциации таежно-степных сосновых, лиственнично-сосновых и лиственничных с участием сосны в комплексе со степными сообществами как демутационно-восстановительные ряды лесных сообществ (рис. 2).
В легенде к карте таежно-степных сообществ (эпиассоциаций) ключевого участка западного побережья оз. Байкал (бассейна р. Таловки, Приольхонье), отражена возрастная структура древостоев ассоциации, состав доминантов и кодоминантов конкретной возрастной группы, наличие или отсутствие подроста и его состав, доминирование или отсутствие такового мелколиственных пород, возобновление в зависимости от фактора воздействий (пожар, рубка, выпас, сенокос), наличие или отсутствие травянистых сообществ демутационно-восстановительного ряда, структурно-динамически и генетически обусловленных составляющих, как единое целое образующих одну эпиассоциацию. Эпиассоциации жестко привязаны к конкретным местоположениям – склонам разных экспозиций и крутизны, водоразделам определенной морфоструктуры, шлейфам склонов разных экспозиций, межгорным распадкам и речной долине.
К примеру, эпиассоциация сосновых с лиственницей и лиственнично-сосновых рододендроновых (рододендрон даурский) с душекией (душекия кустарниковая) брусничных мезофитных лесов на седловинах водоразделов (№ 1, рис. 8) включает березово-сосновый подрост на гари; осиново-березовые леса (возрастная группа) до 40 лет; березово-осиновые леса (возрастная группа до 40 лет); лиственнично-сосновые с березой и осиной без подроста леса (возрастная группа до 40 лет); осиново-лиственничные с березой без подроста леса (возрастная группа до 40 лет); сосновые с березой без подроста леса (возрастная группа до 60 лет); сосновые с березой леса (возрастная группа до 80 лет); сосново-лиственничные с березой и осиной без подроста леса (возрастная группа до 80 лет); сосново-лиственничные с березой и осиной с подростом сосны и лиственницы, березы леса (возрастная группа до 100 лет) и выше. Данная эпиассоциация включает все возможные динамические составляющие лесного сообщества, формирующегося в конкретных условиях – седловина водораздела. Здесь приводится характеристика состава доминантов ярусов, их возрастная структура, характер подроста и возобновления в комплексе со степными сообществами, являющимися демутационно-восстановительными рядами таежно-степных эпиассоциаций.
Эпиассоциация сосновых с лиственницей (№ 6, рис. 8) петрофитных редкотравных с осокой большехвостой, таволгой и участием мхов мезофитные леса на эродированных склонах включает сосновые с лиственницей (возрастная группа древостоя до 40 лет) подростом лиственницы и сосны леса; сосновые с лиственницей (возрастная группа древостоя до 80 лет) с подростом лиственницы и сосны леса; травянистые с доминированием житняка гребенчатого и зопника клубненосного сообщества демутационно-восстановительного ряда сосновых с лиственницей лесов; сосновые с лиственницей (возрастная группа древостоя до 80 лет) с подростом лиственницы, сосны и березы; травянистые с доминированием осоки твердоватой и мятлика кистевидного сообщества демутационного ряда сосновых с лиственницей лесов. Показаны видовой и возрастной составы древостоя, образующих ассоциации, которые в комплексе с травянистыми сообществами образуют эпиассоциацию определенного местообитания. Характер развития отражает тенденции на формирование леса с сокращением участков, занятых травянистыми сообществами. Здесь отмечен подрост сосны вне полога леса.
Эпиассоциация елово-лиственничных с сосной и участием ив (ива – Salix taraikensis и грушанколистная – Salix pyrolifolia) широкотравных (какалия копьевидная – Cacalia hastata, тмин обыкновенный – Carum carvi, купальница азиатская – Trollius asiatica, хвощ луговой – Equisetum pratense, чемерица Лобеля – Veratrum lobelianum) лесов низких надпойменных террас долины р. Таловки включает лиственнично-березовые (береза повислая – Betula pendula) с осиной без подроста леса (возрастная группа до 40 лет); березовые с осиной с участием ив осоковые (осока клопоносная и двуокрашенная – Carex pamirensis subsp. dichroa заболоченные леса (возрастная группа до 60 лет); травянистые с доминированием полевицы монгольской (Agrostis mongolica), лисохвоста лугового, осоки безжилковой – Carex enervis , тмина обыкновенного, чемерицы Лобеля, пушицы рыжеватой, купальницы Кытманова (Trollius kytmanovii) сообщества, используемые в качестве сенокосных угодий; лиственничные с елью и березой с подростом лиственницы и березы и участием ив и смородины черной (Ribes nigrum) условнокоренные для таких местообитаний леса; травянистые с доминированием овсяницы красной –Festuca rubra, полевицы монгольской, мятлика лугового – Poa pratensis, погремка – Rhinanthus vernalis и пушицы рыжеватой заболоченные сообщества, часто используемые как сенокосные угодья и генетически связанные с елово-лиственничными лесами долины Таловки.
Составленная карта таежно-степных эпиассоциаций ключевого участка района исследований – бассейн р. Таловки (рис. 2) включает все динамические и демутационные составляющие лесных и травянистых сообществ, формирующихся в современных контрастных условиях среды, однако генетически связанные и имеющие общую направленность развития. На этом ключевом участке, с домированием сосны и лиственницы в древостое, структура ценозов сложна в аспекте их пространственной неоднородности. В составе эпиассоциации лиственничных зеленомошно-осоково-разнотравных часто присутствует кедр от 2 до 27 лет, при этом отмечается он не только на ключевых участках, но и в структуре растительности всего района исследований. В эпиассоциациях остепненных лиственничников, разных типов их местообитаний, часто существенную роль играют мхи, характерные для полидоминантной темнохвойно-светлохвойной тайги. Здесь развит подрост лиственницы до 17 лет, характерен разновозрастный древостой разной сомкнутости повсеместно, включая более «степную» часть района исследований. Часто 2–17-летние сосны и лиственницы образуют самостоятельные ценозы вне полога древостоев. По возрастному составу превалируют лиственницы от 2 до 30 лет с несколькими деревьями от 60 до 100 лет. Напочвенный покров лиственничных лесов этого района достаточно сложен. Здесь, наряду со «степняками», обычны лугово-лесные (таежные) виды растений в комплексе со мхами, характерными для полидоминантной темнохвойно-светлохвойной тайги и они образуют особую синузиальную структуру сообществ. Такие сообщества рассматриваются как парагенез [11] в структуре растительности и являются стадией восстановительной динамики лесного (таежного) типа растительности в Прибайкалье на данное время.
Динамика таежно-степных сообществ (эпиассоциаций) западного побережья оз. Байкал
Сокращение в последнее время участков, занятых степными сообществами, особенно в зоне контакта лесов и экстразональной степи, подтверждается многолетними наблюдениями за пространственной изменчивостью таежно-степных эпиассоциаций. Наметились тенденции залесения степей, особенно межгрядовых, межсклоновых и выположенных местоположений их формирования. На всей территории исследований происходит активное внедрение мезофитов в степные сообщества, повысилось проективное покрытие и биопродуктивность степных ценозов. Более отчетливо проявляется ярусность напочвенного покрова таежно-степных эпиассоциаций, где на доминирующие позиции выходят лугово-лесные виды растений вне зависимости от типа их местообитаний. В составе таежно-степных эпиассоциаций зоны контакта тайги и степей отмечено пространственное расширение синузий мхов, характерных для темнохвойно-светлохвойной полидоминантной тайги. Особенной чертой пространственных изменений ценоструктуры растительности западного побережья следует отметить тенденции с весьма активным внедрением древесных экобиоморф в степные сообщества, и не только в зоне контакта. При этом образуются разновозрастные куртины от всходов до 25–30-летних деревьев. Ранее были отмечены отдельно стоящие среди степей Pinus sylvestris и Larix sibirica в возрасте 40–50 лет. Процессы общей мезофитизации видового состава и облесения степей отмечены по всему западному побережью оз. Байкал. Генезис растительности региона на протяжении голоцена, современные динамические тенденции сообществ, а также снижение и локализация влияний антропогенных факторов позволяют предположить начало стадии формирования здесь лесного (таежного) типа растительности как особой формы инварианта светлохвойной тайги, характерной только для этого региона Прибайкалья. Это и обусловило пространственную динамику светлохвойных лесов и степных соообществ в аспекте занимаемых ими площадей. По данным исследований последних лет [2] на западном побережье оз. Байкал на протяжении последних 4 тыс. лет происходили достаточно глубокие изменения в составе и структуре растительности региона с периодами расширения лесного комплекса и относительным расширением травянистого на определенных этапах. Изменение соотношения степной и лесной растительности указывает на векторное ослабление аридности климата, но на протяжении последних 2 тыс. лет проявлялись кратковременные этапы ее усиления до 1500, около 900 и 200 лет назад. Вероятно, на двух последних этапах и произошло формирование ксерофитной растительности в Приольхонье, к тому же эти процессы были усилены и воздействием антропогенных факторов.
Таежно-степные сообщества западного побережья оз. Байкал выступают как стадия формирования хвойных лесов на территории прибрежья озера на современном этапе развития растительного покрова и являются индикаторами направленности динамики природной среды региона в целом. Наличие лесных сообществ – соснового и лиственничного древостоя среди степей в центральной части побережья с присутствием растений Bergenia crassifolia, Linnaea borealis, Pyrola rotundifolia, Iris ruthenica, Maianthemum bifolium, характерных для полидоминантной темнохвойно-светлохвойной зональной тайги и наличие синузий мхов, обычных для темнохвойных лесов в составе остепненных, состоящих из Pinus sylvestris, Larix sibirica лесных сообществ, являются показателем своеобразия структуры, динамики и генезиса растительности региона.
В зоне непрерывного контакта лесных и степных сообществ выявились существенные перестройки вертикальной и пространственной их структур. Так, одновременно с формированием в древостоях лесных (таежных) сообществ устойчивого подроста из Pinus sylvestris, Larix sibirica, отмечается активное внедрение древесных экобиоморф в травянистые (степные) сообщества в виде куртин или отдельно стоящих групп подроста до 17 лет. В последние годы в составе травянистых (степных) сообществ, основу которых составляют ксерофиты, такие как Artemisia commutata, Heteropappus altaicus, Phlomis tuberosa, Poa botryoides, Agropyron cristatum, отмечены всходы древесных пород – сосны и лиственницы. В составе напочвенного покрова светлохвойных лесов повсеместно усиливаются позиции лесных видов растений с расширением синузий из Drepanocladus uncinatus, Mnium cuspidatum, Dicranum polysetum, Rhytidium rugosum и Vaccinium vitis-idaea. На доминирующие позиции в травостоях выходят Astragalus versicolor, Galium verum, Aster alpinus, Lupinaster pentaphyllus, Potentilla tanacetifolia, Campanula glomerata, тогда как степные злаки – Festuca lenensis, Koeleria cristata, Poa botryoides, Agropyron cristatum менее обильны и образуют стадию динамики сообществ в течение вегетационного периода. Наличие подроста и всходов сосны и лиственницы в травянистых степных сообществах свидетельствует о тенденциях расширения древесных экобиоморф в связи с увеличением температуры и перераспределение среднегодовых осадков по сезонам года в Прибайкалье в последние 30-50 лет. Вместе с сокращением площадей, занятых степными сообществами, в напочвенном покрове лесов происходит пространственное расширение синузий мхов, характерных для полидоминантной темнохвойно-светлохвойной тайги.
Заключение
Выявленные, методом крупномасштабного картографирования, структурно-динамические особенности растительных сообществ центральной части западного побережья оз. Байкал (Приольхонье) позволяют говорить о процессах формирования тайги на месте остепненных сосновых, редкостойных лиственничных лесов и экстразональных степных сообществ, сформировавшихся на поздних стадиях голоцена под воздействием антропогенных факторов последних десятилетий в регионе. В условиях экстразональности степи западного побережья, где виды растений центрально-азиатской и общеазиатской флор составляют около 1/10 от общего видового состава, можно утверждать, что современные таежно-степные сообщества района исследований есть стадия восстановительной динамики и развитие лесов таежной зоны. Дальнейшее формирование таежно-степных сообществ в аспектах структурных изменений на фоне флуктуаций среднегодовых осадков и среднегодовых зимних температур будет направлено в сторону формирования лесов с образованием небольших, по занимаемой площади, ксрефитно-петрофитных травянистых группировок на каменистых эродированных склонах и по каменистым грядам. Светлохвойные леса района исследований в комплексе со степными сообществами являются, по сути, единым целым в процессе фитоценогенеза. Основным фактором природной среды, обусловившим пространственную стратиграфию растительного покрова района исследований, является местоположение конкретного сообщества в системе гор Западного Прибайкалья. Это находит отражение в структуре растительности, главным образом в подросте, подлеске и напочвенном покрове для лесов и в флористическом составе травянистых (степных) сообществ. Экспозиционность склонов гор определяет только степень сомкнутости древостоев и обилие подроста для лесов и обилие того или иного вида растения в составе травянистых (степных) сообществ. Среди степных участков активно формируются лесные сообщества с достаточно устойчивым возобновлением, зачастую с синузиями мхов, характерных для полидоминантной темнохвойно-светлохвойной тайги. В условиях экстразональности степи, невыраженности горно – степного и горно – лесостепного поясов в районе исследований формируются особые сообщества – «таежно – степные», отражающие парагенез в структуре растительности региона. По характеру развития они являются стадией восстановительной (на разных периодах развития – деструктивной) динамики лесов Прибайкалья и являются выражением сложности структуры и неоднозначности генезиса растительности таежной зоны Прибайкалья.
Библиографическая ссылка
Сизых А.П. КАРТОГРАФИЧЕСКОЕ ИЗУЧЕНИЕ РАСТИТЕЛЬНЫХ СООБЩЕСТВ КОНТАКТА СРЕД // Успехи современного естествознания. 2015. № 1-4. С. 621-634;URL: https://natural-sciences.ru/en/article/view?id=34867 (дата обращения: 03.06.2026).