


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
SUSTAINABLE GROWTH OF PROGENY OF FINNISH SPRUCE AND CHEMICAL COMPOSITION NEEDLES

Для лесных пород актуальна селекция на быстроту и устойчивость роста и важен поиск признаков, связанных с этими показателями. Известны работы, где показаны различия в содержании химических элементов в хвое у клонов сосны обыкновенной; при этом предполагается, что они могут быть связаны с быстротой роста и генетическими отличиями [5]. В работах по ели финской, которая является естественным гибридом ели сибирской и европейской, было обнаружено, что потомства ее популяций с преобладанием признаков ели сибирской накапливают достоверно больше Mn, Ti, P, Cu, Ba, Sr, Pb и при повышении зольности почвы повышают и зольность хвои, в отличие от более «европейских» популяций, которые снижают их [4] и которые имеют повышенную продуктивность потомства [2, 3].
Целью исследования было выяснение влияния химического состава хвои на рост отдельных семей ели финской, выращенных в разных условиях.
Материалы и методы исследования
Изучали потомство ели финской (Picea×fennica (Regel) Kom.), полученное из семян от свободного опыления и выращенное в тест-культурах в крупном опыте [2]. Опыт заложен в 1991 г. в Пермском крае, в кв. 41 Ильинского лесничества. Почва среднесуглинистая, обедненная гумусом в результате сдвига почвы при корчевке (биотоп С2). В тест-культуры (испытательные культуры, ИК) высажено 22,4 тыс. саженцев по схеме 2,5×1,0 м. Сохранность растений составила 74,7 %. Представлены потомства: из 7 естественных популяций от 301 плюсового дерева и из лесных культур (5 участков) от 224 деревьев, в том числе 152 плюсовые и 72 – обычные деревья. Контроль – семена от 30 случайных деревьев от каждой из всех популяций. Посадочный материал выращивали в теплице и в школе. Измерения проведены в 21 год у 16755 растений, в том числе 1435 шт. в контроле.
Потомство этих же деревьев изучали еще на одном участке ИК, заложенном на супеси в биотопе В2, где посадки создавали трехлетними сеянцами на вырубке по раскорчеванным полосам тремя рядами по схеме 0,7×0,7 м, с расстоянием между полосами 8–20 м. Такая схема мало подходила для опыта, но позволила выяснить реакцию семей на конкуренцию. Этот участок, названный «густые культуры», заложен в кв. 5 Юго-Камского лесничества Оханского лесхоза на площади 8 га. Высажено 16,1 тыс. растений на 1711 делянок, потомством 462 деревьев с тем же контролем. В 2010 г. в 23-летнем возрасте деревья были измерены и проведено осветление культур с вырубкой 15 м3/га древесины лиственных пород. Культуры сильно затенялись березой и осиной, которые местами превышали высоту ели на 1–5 м. То есть на этом участке ель испытывала не только конкуренцию от растений своего вида, но и сильное угнетение со стороны лиственных пород. Сохранилось всего 34 % растений, поэтому оставили семьи, где было по 6 растений и более. Выборка на семью составила в среднем 13 шт. и средняя ошибка высоты оказалась 8,4 %. На Ильинском участке измеряли в среднем 29 растений на семью и средняя ошибка высоты была 5,4 %. Парные измерения высот оказались для 392 семей и 21 варианта контроля.
В результате анализа этих пар семей, который будет показан ниже, были выделены 106 семей с разной реакцией на густоту выращивания, от которых в тест-культурах выбирали по одному типичному дереву и брали смешанный образец 1, 2 и 3-летней хвои из средней части кроны при объеме срезанных ветвей 1,0–1,5 л. Хвою для химического анализа брали только на участке с «редкими» культурами. Далее проводили атомно-абсорбционный анализ хвои по 30 элементам [1, 6]. Анализ осуществляли на дифракционном спектрографе СТЭ-1 методом испарения из кратера угольного электрода.
Результаты исследования и их обсуждение
В потомстве естественных популяций парные значения высот 216 семей на обоих участках колебались от 72 до 137 %, и корреляция между высотой семей в редких и загущенных культурах отсутствовала (r = 0,04). Такая же корреляция (r = 0,01) была получена и для 176 семей из популяций культур (рис. 1).
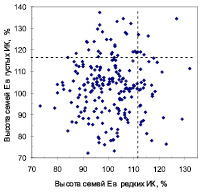
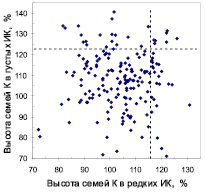
Рис. 1. Поля корреляций между высотами семей в редких и в густых испытательных культурах в потомстве естественных популяций (семьи Е, слева) и в потомстве популяций из культур (семьи К, справа); пунктирные линии отделяют 15 % быстрорастущих семей
Изучение химического состава хвои требовало очень больших затрат, поэтому мы выбрали только контрастные семьи. Чтобы как-то их называть, взяли за основу реакцию семьи на густоту выращивания. Именно густота наиболее радикально влияет на развитие древостоев [3]. Поэтому если при увеличении густоты семья снижала высоту, то мы называли ее «семья-светолюб». Если же она ее увеличивала – это была «семья-конкурентник. Был выделен и третий тип – «семьи-универсалы», но провести их анализ мы не могли по финансовым ограничениям. Для оценки стабильности роста был введен показатель «индекс стабильности роста» (ИСР): отношение высоты семьи в густых ИК к ее высоте в редких. Если ИСР > 1,0 – семья повышает высоту в густых ИК, если меньше 1,0 – снижает ее. Корреляция высот семей с ИСР оказалась слабой (r = 0,14), а значимых ее связей с морфометрией матерей мы не нашли. При такой ситуации и при отнесении к быстрорастущим около 15 % семей в каждом отдельном испытании семей, которые одинаково хорошо растут и в густых, и в редких культурах, становится почти в 5 раз меньше (рис. 1): всего 5 семей из естественных популяций (2,3 %) и 7 семей из культур (4,0 %) могут быть зачислены в таковые (в среднем 3,2 %). При более интенсивном отборе лучших семей, например 10 %-ном, который на рисунках не показан, их может быть всего лишь 1–2 %.
Далее мы выбрали контрастные по росту семьи. В редких ИК было выбрано 55 семей из естественных популяций и 51 семья из популяций культур, из которых парные высоты в густых ИК оказались для 44 и 47 семей соответственно (рис. 2).
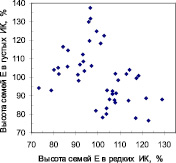
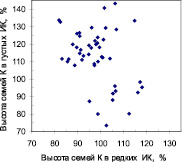
Рис. 2. «Семьи-конкурентники» (кластер точек слева вверху) и «семьи-светолюбы» (точки справа внизу) в потомстве естественных популяций (семьи Е) и в потомстве культур (семьи К)
Далее в редких тест-культурах выбирали по одному типичному дереву от семьи для химического анализа хвои. В результате атомно-абсорбционного анализа хвои полные ряды без нулевых значений были получены по 12 химическим элементам (ХЭ): Ni, Cr, Mn, V, Ti, P, Cu, Zn, Ba, Sr, Zr, Pb. Их сравнение показало, что в потомстве естественных популяций (Е) более активно накапливают ХЭ семьи-конкурентники. Сумма концентраций всех 12 элементов у них оказалась в 1,28 раза выше, чем у семей-светолюбов, и по 8 микроэлементам из 12 повышение было достоверно; особенно значимое повышение (в 1,39–1,76 раза) отмечено по Ti, Mn и Zn (таблица). Следует отметить, что в сумму концентраций ХЭ наибольший вклад вносят марганец и фосфор и доля их составляет 37–42 % [4]. Поэтому именно марганец повлиял на эту сумму наиболее радикально.
Однако в потомстве культур такого согласованного повышения не наблюдалось. Всего лишь три элемента из 12 увеличили концентрации в семьях-конкурентниках: никель в 1,57, марганец в 1,19 и цирконий – в 1,4 раза. По остальным девяти элементам повышение было недостоверно. По зольности хвои различия между КС и КК были недостоверны, тогда как в потомстве Е повышение зольности хвои у конкурентников в 1,08 раза оказалось достоверным (таблица).
Полученные данные о химическом составе хвои позволяют предполагать существование неких физиологических различий между потомствами этих групп популяций. Семьи-конкурентники из естественных популяций, видимо, формируют более морозоустойчивую хвою, накапливая в ней в 1,28 раза больше химических элементов и повышая ее зольность в 1,08 раза. В потомствах культур, которые по своей морфологии тяготеют к ели европейской [2], семьи делают это менее активно и по большинству химических элементов их повышения недостоверны.
Далее мы проанализировали влияние суммы концентраций химических элементов хвои на рост семей в разных тест-культурах. Рассмотрим два поля корреляции, поместив их рядом: для случая редких культур слева, а для густых – справа (рис. 3).
Содержание химических элементов в хвое ели финской с разной реакцией семей на конкуренцию
|
Статистический показатель |
Индекс стабильности роста |
Содержание химических элементов в хвое, мг/кг |
Зольность, % |
||||||||||||
|
Ni |
Cr |
Mn |
V |
Ti |
P |
Cu |
Zn |
Pb |
Ba |
Sr |
Zr |
Сумма |
|||
|
Естественные популяции, светолюбы (ЕС), n = 30 |
|||||||||||||||
|
Среднее |
0,796 |
1,53 |
0,58 |
376,0 |
0,58 |
4,00 |
526,5 |
1,32 |
15,03 |
0,26 |
169,1 |
60,03 |
1,32 |
1156 |
5,82 |
|
Стандартное отклонение |
0,097 |
0,682 |
0,059 |
189,72 |
0,059 |
1,94 |
67,03 |
0,446 |
8,07 |
0,344 |
58,04 |
20,53 |
0,65 |
241,0 |
0,616 |
|
Коэффициент вариации, % |
12,2 |
45 |
10 |
50 |
10 |
48 |
13 |
34 |
54 |
132 |
34 |
34 |
49 |
21 |
11 |
|
Естественные популяции, конкурентники (ЕК), n = 25 |
|||||||||||||||
|
Среднее |
1,248 |
1,39 |
0,63 |
622,16 |
0,63 |
7,05 |
563,6 |
1,34 |
20,84 |
0,17 |
181,0 |
76,26 |
1,63 |
1477 |
6,27 |
|
Стандартное отклонение |
0,124 |
0,447 |
0,083 |
84,83 |
0,083 |
2,34 |
83,54 |
0,545 |
8,35 |
0,209 |
72,55 |
27,31 |
0,61 |
231,1 |
0,827 |
|
Коэффициент вариации, % |
9,9 |
32 |
13 |
14 |
13 |
33 |
15 |
41 |
40 |
123 |
40 |
36 |
37 |
16 |
13 |
|
Отношение ЕК/ЕС |
0,91 |
1,08 |
1,65 |
1,08 |
1,76 |
1,07 |
1,02 |
1,39 |
0,65 |
1,07 |
1,27 |
1,24 |
1,28 |
1,08 |
|
|
tэксп |
–0,90 |
2,48* |
6,27* |
2,48* |
5,09* |
1,76 |
0,14 |
2,56* |
–1,19 |
0,65 |
2,40* |
1,79 |
4,93* |
2,20* |
|
|
Культуры, светолюбы (КС), n = 11 |
|||||||||||||||
|
Среднее |
0,826 |
1,01 |
0,51 |
413,3 |
0,51 |
5,27 |
483,3 |
1,58 |
15,37 |
0,36 |
125,8 |
54,1 |
1,17 |
1102,2 |
5,1 |
|
Стандартное отклонение |
0,068 |
0,23 |
0,09 |
134,1 |
0,09 |
0,99 |
80,37 |
0,47 |
8,48 |
0,33 |
56,03 |
26,42 |
0,39 |
187,6 |
0,90 |
|
Коэффициент вариации, % |
8,2 |
23,2 |
17,8 |
32,5 |
17,8 |
18,7 |
16,6 |
29,7 |
55,2 |
92,3 |
44,5 |
48,8 |
33,5 |
17,0 |
17,8 |
|
Культуры, конкурентники (КК), n = 40 |
|||||||||||||||
|
Среднее |
1,267 |
1,59 |
0,54 |
493,3 |
0,54 |
5,65 |
507,0 |
0,87 |
15,62 |
0,21 |
128,5 |
52,7 |
1,64 |
1208,1 |
5,4 |
|
Стандартное отклонение |
0,124 |
0,61 |
0,07 |
127,5 |
0,07 |
2,00 |
73,28 |
0,59 |
11,71 |
0,17 |
46,55 |
20,01 |
0,58 |
207,5 |
0,73 |
|
Коэффициент вариации, % |
9,8 |
38,3 |
13,5 |
25,8 |
13,5 |
35,5 |
14,5 |
68,1 |
75,0 |
78,3 |
36,2 |
38,0 |
35,1 |
17,2 |
13,5 |
|
Отношение КК/КС |
1,57 |
1,07 |
1,19 |
1,07 |
1,07 |
1,05 |
0,55 |
1,02 |
0,59 |
1,02 |
0,97 |
1,40 |
1,10 |
1,06 |
|
|
tэксп |
4,80* |
1,17 |
1,77 |
1,17 |
0,87 |
0,88 |
4,19* |
0,08 |
–1,41 |
0,15 |
–0,17 |
3,12* |
1,62 |
1,01 |
|
Примечание. Е – естественные популяции, К – культуры; * – различие между средними достоверно при P < 0,05; n – число семей.
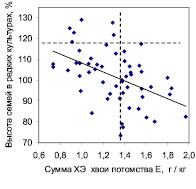
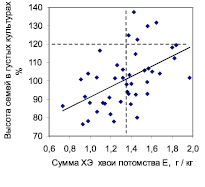
Рис. 3. Влияние суммы концентраций химических элементов в хвое на рост семей в редких и в густых тест-культурах в потомстве естественных популяций
На участке редких культур, где брали образцы хвои, т.е. на левом графике, связь между суммой концентраций ХЭ и ростом семей была отрицательна (r = –0,47 ± 0,10), тогда как в густых культурах на правом графике (где хвою не изучали) она стала положительной (r = 0,51 ± 0,11). Так как хвою на анализ из густых культур мы не брали, то объяснить влияние суммы концентраций ХЭ в ней на высоту семей там, где эта хвоя не росла и «не работала», можно только тем, что этот химический показатель как-то связан с генетическими особенностями семьи и конкретного дерева. Поэтому можно полагать, что сумма концентраций химических элементов хвои проявляет себя как некий химический признак (хемопризнак) дерева, увеличение которого снижает высоту деревьев в редких культурах и, наоборот, увеличивает ее в густых культурах.
Для оценки возможного практического эффекта использования данного хемопризнака можно имитировать отбор семей по сумме концентраций ХЭ, ориентируясь на значение 1,35 г/кг, и отбирать в редких культурах семьи с малым, а в густых культурах – с большим содержанием суммы концентраций изученных ХЭ в хвое (вертикальная линия на графиках). После этого частота быстрорастущих семей (горизонтальная линия на графиках) составит на левом графике 5/27 = 0,185 и по отношению к исходной их частоте, равной 5/55 = 0,091, новая частота окажется выше в 2,03 раза. На правом графике исходная их частота была 5/44 = 0,114, а новая оказалась 5/24 = 0,208, что выше исходной в 1,82 раза. Отобранные по указанным критериям семьи имеют среднюю высоту в редких культурах 108,4 ± 2,2 % и в густых 107,2 ± 3,1 %. Это неплохой результат с учетом повышения частоты лучших семей в 1,8–2 раза. Если учесть, что для получения образцов хвои потомство можно выращивать всего 3–4 года, то при отсутствии взаимодействий «генотип-возраст» по изучаемым признакам возможна ранняя диагностика нужных семей, что необходимо проверить.
Сумма концентраций ХЭ хвои коррелирует также с индексом стабильности роста (r = 0,60 ± 0,10). Поэтому если мы отбираем семьи-конкурентники, то их нужно искать среди семей с суммой концентраций ХЭ выше среднего. Но если мы селекционируем семьи-светолюбы, то их следует искать среди семей, накапливающих химические элементы на низком уровне. Химические показатели хвои влияют на рост потомства более сильно, чем морфометрические показатели – высота и сбег ствола, масса семени и шишек; ранее в результате использования последних нам удалось повысить число лучших семей в 1,4–1,7 раза и высоту потомства лишь на 3,0 % [3].
Дальнейшие исследования должны дать ответы на три вопроса:
а) насколько устойчивы во времени различия между семьями по составу химических элементов;
б) в каком возрасте семей хвоя приобретает отмеченные выше отличия;
в) какие химические элементы маркируют быстрорастущие семьи.
Выводы
1. Частота быстрорастущих семей (15 % семей в каждом испытании), устойчиво растущих и в густых, и в редких культурах, оказывается 3,2 %. Поэтому для отбора 50 матерей для сорта-популяции нужно испытать не менее 1,6 тыс. потомств в разных условиях. В связи с этим на первом этапе селекции усилия следует направить на сорта для выращивания, например, только на плантациях с редким размещением деревьев.
2. Данные о химическом составе хвои показали, что семьи-конкурентники из естественных популяций ели финской с большей долей форм, близких к ели сибирской, накапливают в хвое в 1,28 раза больше химических элементов с повышением ее зольности в 1,08 раза. По-видимому, это связано с их большей морозоустойчивостью. В потомствах культур, которые по морфологическим признакам уклоняются в сторону ели европейской, семьи делают это менее активно и повышение их концентраций недостоверно. Это позволяет рассматривать химический состав хвои как отражение физиологических и генетических отличий между изученными группами популяций.
3. Влияние суммы концентраций химических элементов хвои на рост семей в редких и густых тест-культурах оказалось разнонаправленным. В редких культурах, где брали образцы хвои, связь между суммой концентраций химических элементов и ростом семей была отрицательна (r = –0,47 ± 0,10), тогда как в густых культурах, где хвою не изучали, она оказалась положительной (r = 0,51 ± 0,11). В связи с этим можно полагать, что сумма концентраций химических элементов хвои обусловлена генетически и при ее увеличении высота потомства в редких культурах снижается, а в густых культурах, наоборот, возрастает.
4. Имитация отбора семей по сумме концентраций химических элементов с ориентацией на значение 1,35 г/кг показала увеличение высоты семей в редких культурах до 108,4 ± 2,2 % и в густых до 107,2 ± 3,1 % от контроля, с повышением частоты лучших семей в 1,8–2 раза.
Работа выполнена при частичной финансовой поддержке задания 2014/153 государственных работ в сфере научной деятельности в рамках базовой части госзадания Минобрнауки России, проект 144 № ГР 01201461915.
Библиографическая ссылка
Рогозин М.В., Жекина Н.В. УСТОЙЧИВЫЙ РОСТ ПОТОМСТВА ЕЛИ ФИНСКОЙ И ХИМИЧЕСКИЙ СОСТАВ ХВОИ // Успехи современного естествознания. 2016. № 5. С. 79-84;URL: https://natural-sciences.ru/en/article/view?id=35902 (дата обращения: 02.07.2026).