


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
BIOLOGICAL PRODUCTIVITY AND CONSERVATION OF NATURAL PLANT COMMUNITIES STEPPES OF KHAKASSIA IN LONG TERM OPERATION

Степи Хакасии расположены в основном на юге Минусинской котловины. По классификации Е.М. Лавренко [6] приуроченность степей к южным черноземам позволяет отнести эти травяные экосистемы к настоящим засушливым степям, растительный покров которых состоит из крупнодерновинных и мелкодерновинных злаков, а также из корневищных многолетников. Заметное участие в травяных экосистемах настоящих степей принимают кустарники и виды ксерофитного разнотравья. В настоящее время, когда хозяйственная деятельность усиливается, познание этих вопросов в рамках всех ритмов природных явлений, особенно с учетом климатических изменений, представляется очень актуальным.
Цель исследования – изучить структуру и динамику растительного вещества степных ландшафтов Хакасии юга Красноярского края, на основе многолетних стационарных работ, с позиций системного подхода, этапы которых увязаны между собой единой идеей.
Материалы и методы исследования
Детальные исследования проводились в фациях, до и после снятия заповедного режима на экспериментальном Новониколаевском физико-географическом стационаре, расположенном в Койбальской степи Минусинской котловины на юге Красноярского края. Степной стационар находится между горными системами Западного, Восточного Саяна и Кузнецкого Алатау с мелкогрядовым и плоскоравнинными формами рельефа. Абсолютные высоты составляют 300–500 м. Климат котловины довольно теплый: средняя температура июля составляет 19 °С, среднегодовая положительна 0,9 °С, выпадает 350–450 мм осадков в год, максимальное количество приходится на летние месяцы (50–60 %). Вегетационный период длится с середины апреля до середины октября.
Стационарные исследования в заповедных условиях велись в 1970–1994 гг. Результаты коллективного труда за эти годы исследований освещены в многочисленных публикациях и монографиях [1, 8]. С 1994 г. на полигон-трансекте снят заповедный режим, с 2000 г. наблюдения по продуктивности продолжены в условиях естественного состояния фаций. Результаты наблюдений опубликованы в коллективных монографиях [3, 10].
Основные понятия и термины по биологической продуктивности изложены в ряде работ [7, 9]. Биологическая продуктивность характеризуется запасом надземных органов растений, полученных на единицу площади, которая делится на следующие фракции: зеленая масса, ветошь и подстилка. Зеленую массу разбирали по видам и делили на жизненные формы. Из состава степной флоры для анализа были взяты растения, играющие наиболее выраженную роль в сложении травостоя сообществ – эдификаторы и доминанты, а также некоторые виды растений, хотя и не играющие большой фитоценотической роли, но имеющие интересные биологические особенности. Все пробы растительного вещества высушивались в течение 8–12 часов при температуре 105 °С до абсолютно сухого веса, затем взвешивались. Ошибка средней для данных надземной массы составила ±10–16 %.
Результаты исследования и их обсуждение
Койбальская степь включает в себя разные по характеру сообщества, различающиеся в зависимости от условий мезорельефа, проективного покрытия, высоты травостоя и видового разнообразия. Значительную часть продуктивности сообществ обеспечивают доминирующие виды, которые объединены в группы по жизненным формам. Изучение жизненных форм способствует более глубокому познанию структуры и динамики растительных сообществ, а распределение удобнее рассматривать по элементам рельефа (таблица).
Характеристика показателей растительных сообществ в условиях заповедного и естественного режима Койбальской степи с 1970 по 2015 г.
|
Элемент мезорельефа |
Транзитная часть южного склона |
Транзитная часть северного склона |
|
Почва |
Чернозем южный среднегумусный среднемощный солонцеватый |
Чернозем южный малогумусный среднемощный карбонатный солонцеватый |
|
Растительное сообщество |
Мелкодерновинно-злаково-ковыльное с караганой |
Разнотравно-овсецово-ковыльное с караганой |
|
Эдификаторы, доминанты, и другие виды растений травяного покрова |
Stipa Krylovii, Helictotrihom desertorum Festuca calesiaca, Koeleria cristata, Cleistogenes squarrosa, Carex pediformis, Artemisia frigida, Caragana pygmaea, Aneurolepidium pseudoagropyrum |
Helictotrihom desertorum, Stipa Krylovii, Festuca valesiaca, Koeleria cristata, Cleistogenes squarrosa, Carex duriuscula, Artemisia frigida, Caragana pygmaea, Poa stepposa |
|
Заповедный режим |
||
|
Количество видов на 1 м2 |
14–17 |
11–23 |
|
Высота травостоя, см |
10–65 |
10–60 |
|
Проективное покрытие, % |
60–70 |
80–90 |
|
Группы растений: Злаки, % |
54–80 |
45–67 |
|
Осоки, % |
4–18 |
12–21 |
|
Разнотравье, % |
2–10 |
9–25 |
|
Полукустарнички, % |
1–8 |
1–9 |
|
Кустарники, % |
8–13 |
8–15 |
|
Естественный режим |
||
|
Количество видов на 0,25 м2 |
5–6 |
7–12 |
|
Высота травостоя, см |
10–70 |
10–60 |
|
Проективное покрытие, % |
50–60 |
50–70 |
|
Группы растений: Злаки, % |
35–80 |
47–67 |
|
Осоки, % |
2–16 |
2–30 |
|
Разнотравье, % |
4–23 |
13–33 |
|
Полукустарнички, % |
1–40 |
3–8 |
|
Кустарники, % |
4–25 |
1–13 |
Южные склоны характеризуются значительной инсоляцией, прогреваемостью почвы, недостаточным атмосферным увлажнением. Почвенный покров неоднородный, представлен черноземами южными маломощными среднегумусными солонцеватыми. Ниже по склону напряженность абиотических факторов постепенно ослабевает (угол наклона 5–7°), транзитные процессы аккумуляции и сноса вещества уравновешены, почвы маломощны и щебнисты, но содержание гумуса увеличивается [8].
Растительный покров в транзитной части полигон-трансекта склона южной экспозиции представлен мелкодерновинно-злаково-ковыльным сообществом. Эти сообщества относятся к переходным от мелкодерновинно-злаковой группы к крупнодерновинной злаковой [5]. С первыми их сближает полидоминантная структура с господством дерновинных злаков. Для них характерны виды: Helictotrihom desertorum, Stipa capillata, Carex pediformis, Aster alpinus, Bupleurum scorzonerifolium, Artemisia glauca, Poa botryoides. Со вторыми – эдификаторная роль ковыля Крылова и мелкодерновинные виды: Stipa Krylovii, Festuca valesiaca, Koeleria cristata, Poa botryoides, Agropyron cristatum, Carex duriuscula. В местах с близким залеганием материнских пород на слабо сформированных щебнистых почвах превалируют корневищные виды: кустарник – Caragana pygmaea и полукустарничек – Artemisia frigida. Это два вида с вегетативным и семенным типом размножения, с большой продолжительностью и устойчивостью жизни в степных сообществах [4]. Функции видов в сообществе строго распределены: одни – эдификаторы со стабильным доминированием, это Stipa Krylovii и Cleistogenes squarrosa; другие испытывают флуктуации – Festuca valesiaca Koeleria cristata; третьи уходят из фитоценоза при сильном выпасе – Helictotrihom desertorum; четвертые усиливают свои позиции с увеличением солонцеватости – Carex duriuscula.
Долевое участие жизненных форм в мелкодерновинно-злаково-ковыльном сообществе представлено на рис. 1, где показана сравнительная оценка средних показателей: a – заповедного; б – естественного режима.
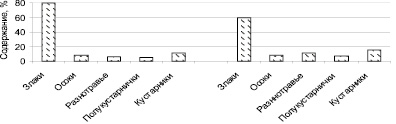
Рис. 1. Содержание жизненных форм в мелкодерновинно-злаково-ковыльном сообществе на южном склоне полигон-трансекта: а – с заповедным режимом (1970–1992 гг.); б – естественным режимом (2000–2015 гг.)
Изучение жизненных форм на южном склоне полигон-трансекта заповедного режима мелкодерновинно-злаково-ковыльного сообщества показало преобладание злаков, которые колеблются от 54 до 80 %, где первое место принадлежит крупнодерновинным злакам: Helictotrihom desertorum, Stipa capillata. Низким содержанием в заповедном режиме отмечены: осоки, разнотравье и полукустарнички до 10 % [2]. Осоки ведут себя по-разному, если в почве нет признаков солонцеватости, доминирует дерновинная форма – Carex pediformis, а на солонцеватых почвах корневищная форма – Carex duriuscula. Смена эдификаторов сообщества после прекращения заповедного режима сопровождается снижением мелкодерновинных злаков: Festuca valesiaca, Koeleria cristata. В отдельные годы они остаются в числе доминирующих видов.
В мелкодерновинно-злаково-ковыльном сообществе естественного режима, по сравнению с заповедными условиями, существование травяного покрова изменено. Проективное покрытие составляет 50–60 %, а содержание крупнодерновинных злаков, Helictotrihom desertorum, Stipa Krylovii, в сухие годы составляет 35 %, а во влажные увеличивается до 80 % (табл. 1). Обилие Festuca valesiaca, Cleistogenes squarrosa, по сравнению с заповедными условиями, так же стабильно увеличивается, но оно может меняться от сочетания тепла и влаги. В травостое с естественным режимом преобладает обилие вида Carex pediformis, при этом снижается обилие Carex duriuscula. Из группы разнотравья уменьшается обилие видов Potentilla acaulis, Galium verum, Dianthus versicolor. В естественном режиме, по сравнению с заповедными условиями, жизненная форма полукустарнички (Artemisia frigida, Thymus minussinensis) увеличивает свое содержание в отдельные годы до 40 %. Выпадающие осадки могут длительное время удерживаться в почве, что позволяет полукустарничкам постепенно расходовать почвенную влагу и накапливать свою массу. В эти годы полукустарнички выступают в роли субдоминанта, а в основном среднее содержание группы полукустарничков до 7 %, независимо от того в каком режиме природной среды они находятся. В современных условиях естественного режима заметно увеличилось долевое участие кустарника Caragana pygmaea – в 2,5 раза по сравнению с заповедными условиями.
Северный склон северной гряды крутой. В денудационной части угол наклона достигает 25°. Рыхлый покров на этом участке склона представлен мелкоземом мощностью не более 0,5 м с редкой дресвой и щебнем кремнистых известняков. Почвы южные щебнистые [1]. Ветровая и радиационная затененность, большая крутизна верхней части северного склона создает особые экологические условия, отличающиеся от южных склонов. Интересной особенностью крутых северных склонов являются процессы новации, связанные с накоплением и длительным сохранением больших запасов снега. В результате медленного таяния в рельефе формируются отрицательные формы – нивальные ниши. В зависимости от степени сформированности нивальных ниш их растительность проходит все стадии первичной сукцессии от луговых микрогруппировок до кустарниковой и древесной [8]. Крутые склоны северной экспозиции заняты разнотравно-овсецово-ковыльным травостоем. Горизонтальное сложение этого сообщества комплексное. На общем фоне пятнами выделяются осочково-вострецовые сообщества со значительным участием полыни холодной – Artemisia frigida, типчака – Festuca и тонконога гребенчатого – Koeleria cristata. В сообществах северного склона на щебнистых почвах произрастает овсец пустынный – Helictotrihom desertorum, который выступает как субдоминант. На рис. 2 показана сравнительная характеристика средних показателей долевого участия жизненных форм при a – заповедном; б – естественном режиме.
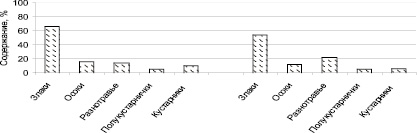
Рис. 2. Содержание жизненных форм в разнотравно-овсецово-ковыльном сообществе на северном склоне полигон-трансекта: а – с заповедным (1970–1994 гг.); б – естественным режимом (2000–2015 гг.)
Одной из характерных черт состава разнотравно-овсецово-ковыльного сообщества является преобладание злаков, которые колеблются от 45 до 67 %, где большая часть принадлежит крупнодерновинным злакам Helictotrihom desertorum, Stipa Krylovii. Сравнение современных условий с заповедным режимом показало, что содержание крупнодерновинных злаков уменьшилось. Все остальные группы жизненных форм ниже 20 %, при этом выделяется группа степного разнотравья в условиях естественного режима, которая увеличилась до 22 %, Увеличение запасов растительного вещества в благоприятные годы происходит в основном за счет этой группы. Значительное снижение содержания кустарников, особенно – Caragana pygmaea, происходит за счет влияния весенних палов.
Продуктивность является одним из важнейших результирующих показателей жизнедеятельности и развития сообщества [6]. Для выяснения экологических особенностей запасов надземной массы крупнодерновинных и мелкодерновинно-злаково-разнотравных сообществ Минусинской степи были построены гистограммы по средним значениям заповедного (1970–1994 гг.) и естественного (2000–2015 гг.) режимов полигон-трансекта. С изменением режима увлажнения и теплообеспеченности из года в год меняется величина запасов фитомассы, а недостаток влаги в корнеобитаемом слое почвы и частые весенние палы в естественном режиме еще больше приводят к уменьшению мортмассы, т.е. ветоши и подстилки (рис. 3).

Рис. 3. Средние запасы надземной массы растительного вещества на полигон-трансекте Минусинской степи при разных режимах, г/м2
Запасы надземной массы в растительных сообществах имеют свои особенности, в зависимости от использования. Общее количество зеленой массы на разных склонах северной и южной экспозиции за годы наблюдений (1970–1994 гг.) при заповедных условиях показали значительное варьирование в сухие и влажные годы (от 80 до 345 г/м2). Зависимость накопления зеленой массы определяется не только с количеством выпавших в данный год осадков, но и с преобразованиями этой влаги в почве и ее запасами с осени. На северном склоне запасы зеленой массы колеблются от 24 до 200 г/м2, ее увеличение происходит за счет накопления влаги в почве из-за медленного таяния снежного покрова. В условиях естественного режима на южном склоне (2000–2015 гг.) запасы зеленой массы колеблются от 96 до 309 г/м2. На северном склоне в разнотравно-овсецово-ковыльном сообществе естественного режима, в зависимости от содержания тепла и влаги и техногенного влияния варьирование значительно ниже – от 9 до 91 г/м2. Данные по максимальным запасам зеленой массы заповедных условий в 3,5 раза выше естественных.
Анализ мортмассы по средним показателям в разных экологических условиях показал существенную разницу ее запасов в заповедных условиях (1970–1994 гг.), имеющих большую амплитуду колебаний – от 30 до 658 г/м2. Запасы мортмассы при естественном режиме (2000–2015 гг.) колеблются от 17 до 153 г/м2. Количество ветоши в заповедных условиях выше в 4,5 раза, а подстилки в 7 раз, по сравнению с естественным режимом (рис. 3). Для Хакасских степей типична своя система динамического равновесия растительного вещества в зависимости от соотношения тепла и влаги и техногенных нагрузок.
Выводы
Полученные результаты по составу жизненных форм и продуктивности надземной массы растительных сообществ Минусинской котловины за длительный период с 1970 по 2015 гг. стационарных наблюдений с разными режимами позволили сделать общие выводы:
1. Результаты сравнительного анализа продуктивности Минусинских степей позволяют понять картину утраченного и выявить тенденции развития степных сообществ в многолетнем режиме функционирования. Система центральноазиатских типов степей подтверждается в основном содержанием видов жизненных форм растительности и продуктивностью фитомассы.
2. Показана роль злаков растительных сообществ как одного из критериев отнесения сообществ к типу настоящих степей. Значительная роль принадлежит крупнодерновинным злакам: Helictotrihom desertorum, Stipa Krylovii, и они занимают лидирующее положение. При смене режимных условий мелкодерновинные злаки Festuca valesiaca, Koeleria cristata также остаются в числе доминирующих.
3. Выявлена устойчивость сообществ, которая обеспечивается в многолетних циклах флуктуациями видов травяного покрова. Установлена продуктивность растительных сообществ при разных режимах: запасы зеленой массы в заповедных условиях колеблются от 80 до 345 г/м2, мортмассы – от 30 до 658 г/м2. Растительные сообщества при естественном состоянии характеризуются запасами мортмассы в 5–7 раз ниже, по сравнению с заповедными условиями. Для Хакасских степей типична своя система динамического равновесия растительного вещества в зависимости от соотношения тепла и влаги и техногенных нагрузок.
4. Стихийные периодически возникающие весенние палы охватывают большие площади, ведут к иссушению местообитания и ксерофитизации растительного покрова. При засухе выявлена высокая устойчивость полукустарничков – полыни холодной – Artemisia frigida и некоторых видов разнотравья – Allium odorum и Galium verum. Отмечено увеличение кустарника – Caragana pygmaea. Во время весенних пожаров происходит угнетение или уничтожение семян ценных кормовых злаков. В этой связи для сохранения банка семян и качества природной среды возникает необходимость пересмотра тактических принципов природопользования в условиях степных ландшафтов.
Библиографическая ссылка
Дубынина С.С. БИОЛОГИЧЕСКАЯ ПРОДУКТИВНОСТЬ ЗАПОВЕДНЫХ И ЕСТЕСТВЕННЫХ РАСТИТЕЛЬНЫХ СООБЩЕСТВ СТЕПЕЙ ХАКАСИИ В МНОГОЛЕТНЕМ РЕЖИМЕ ФУНКЦИОНИРОВАНИЯ // Успехи современного естествознания. 2016. № 5. С. 135-140;URL: https://natural-sciences.ru/en/article/view?id=35912 (дата обращения: 02.07.2026).