


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
CHARACTERISTICS SEED GERMINATION AND SEEDAGE OF PRITOBOLIE WESTERN SIBERIA OF CALLUNA VULGARIS (L.) HULL

Одним из характерных видов, доминирующим в составе травяно-кустарничкового яруса сосновых лесов на суховатых песчаных почвах надпойменных террас рек Притоболья и Русской равнины, а также открытых местообитаний («верещатников») Приатлантики в Европе, является вереск обыкновенный (Сalluna vulgaris).
В настоящее время ареал, морфология, аутэкология и физиология особей вереска изучены достаточно широко. Однако экологические факторы и особенности семеношения и семенного возобновления его ценопопуляций под влиянием пожаров как главного пускового фактора («триггера») возобновления рассмотрены лишь в немногих работах европейских авторов (Whittaker, Gimingham, 1960; Gimingham, 1975).
Вереск успешно размножается и возобновляется вегетативно путем укоренения плагиотропных ветвей в подстилке, а также ежегодно продуцирует огромное количество семян. По исследованиям в Шотландии (Gimingham, 1960), в почвенном банке вересковых ценозов содержится около 158000 всхожих семян на 1 м2. Показано, что на обилие, качество формирующихся семян C. vulgaris и их прорастание влияют тип субстрата, влажность приземного воздуха и освещенность (Leyton, 1955; Gimingham, 1960). Снижение ФАР под пологом древостоев более чем на 40 % приводит к снижению семеношения C. vulgaris и его вытеснению, а максимальное изобилие вереска достигается лишь на открытых участках (Bjorkman, 1945).
В Западной Сибири вереск тесно ассоциирован с сосновыми борами-зеленомошниками и, как и сосна обыкновенная, является типичным видом-пирофитом, адаптированным к пирогенной среде (Санников, 2013). Вспышки его семенного возобновления происходят после уничтожения покрова зеленых мхов с доминированием Pleurozium schreberi (Brid.) Willd. Однако факторы и степень отрицательного влияния этих мхов на прорастание семян и укоренение всходов вереска не изучены.
Целью настоящего сообщения является экспериментальное изучение особенностей семенной репродукции и влияния различных типов напочвенного субстрата на начальные, решающие фазы естественного возобновления ценопопуляций C. vulgaris в сосновых лесах Притоболья.
Материалы и методы исследования
Сравнительный анализ параметров семян и плодов C. vulgaris проведен в популяциях, расположенных на географической трансекте ориентированной с запада на восток ареала: Брянск – Воронеж – Череповец – Нижний Новгород – Красный Октябрь – Заводоуспенское – Курган. Все популяции произрастают под пологом географически замещающих типов сосняков бруснично-вересково-зеленомошных.
Для определения обилия почвенного банка семян вереска заложены две пробные площади в сосняке бруснично-вересково-зеленомошном в подзоне северной лесостепи (Курганская область) под пологом и на открытой вырубке. На каждой из них взяты почвенные высечки размером 10×10×10 см в трехкратной повторности, в лабораторных условиях тщательно разобранные под бинокуляром (Carl Zeiss Stemi) с целью определения банка семян вереска. В конце вегетационного периода на этих же пробных площадях с куртин вереска срезаны побеги с коробочками для подсчета числа семян.
С целью проверки гипотезы о стимулирующей роли пожара на прорастание семян C. vulgaris также взяты высечки в двукратной повторности почвенного субстрата размером 10×10×10 см. Термическое воздействие проведено в двух вариантах:
1) субстрат с полностью выжженной подстилкой;
2) субстрат, прогретый в сухожарном шкафу (ШСП 0.25-60) при температуре 80 °С в течение 15 минут (Wallace, 1917; Elliott 1953; Fenton, 1951). После него модельные высечки высаживались в однородных условиях среды.
Кроме того, в климатической камере (Binder KBF 240) (температура 21 °С, влажность 50 %, длина светового дня не менее 8 часов) заложен контрольный вариант (100 семян).
С целью выявления влияния выжигания мохового покрова (пирогенного обнажения поверхности почвы) на прорастание семян вереска сопоставлены два варианта напочвенного субстрата. Свежеобожженную почвенную высечку и почву с покровом слоя мха Pleurozium Schreberi толщиной 2,5 см высушивали до полной стабилизации массы, а затем помещали равными навесками в чашки Петри и задавали влажность не более 30 % (по отношению к сухой массе). Для исключения влияния ФАР на прорастание семян высечки мха раскладывали в затемненные чашки. Опыт поставлен в трехкратной повторности (50 семян×3).
Для определения объемной влажности субстрата образцы почвы отбирали алюминиевыми бюксами (предварительно определенной массы) с глубины 10–15 см и взвешивали их на электронных весах в лесу, а после высушивания при 105 °С (в течение 6 часов) определяли их объемную влажность. Освещенность под пологом древостоев определяли люкс-метром ТКА-ПКМ.
Возраст кустов вереска определяли по числу годичных колец на дисках поперечных срезов у основания лидирующих по длине побегов.
Элементарная статистическая обработка данных проведена на базе пакета программ Statistica 8.0.
Рис. 1. Карта-схема расположения ценопопуляций выборок C. vulgaris. Примечание: Бр – Брянск, Чп – Череповец, Вр – Воронеж, НН – Нижний Новгород, Тг – Тугулым, Зус – Заводоуспенское, КО – Красный Октябрь, Кг – Курган
Результаты исследования и их обсуждение
Экология произрастания и семеношение. На территории Западной Сибири популяции вереска в большинстве своем сосредоточены по краю стены соснового леса и формируют небольшие группы на открытых участках, не удаляясь далеко от затеняющих их деревьев Pinus sylvestris L. На пробных площадях в типах леса «сосняк бруснично-вересково-зеленомошный» в северной лесостепи Зауралья, на открытом местообитании вереск формирует компактные кусты высотой до 50–60 см. Под пологом леса их средняя высота не превышает 19,6 ± 1,8 см, а диаметр куртин – 2,7 ± 0,7×1,9 ± 0,6 м. Максимальный возраст кустарников изменяется от 45 до 50 лет, что соответствует давности пожара, прошедшего в сосняке. Облиственность годичных побегов, под пологом и на открытых участках, не превышает 4 года.
Число семенных коробочек на одном годичном побеге куста C. vulgaris под пологом древостоя сосны составило 52,04 ± 9,8 шт. (расчетное число семян – 388,03 ± 110,7 шт.), а на открытом месте – 416,34 ± 78,02 шт. коробочек и 3109,04 ± 885,9 шт. семян. Максимальное число коробочек с одного куста C. vulgaris, произрастающего под пологом леса, составило 594 шт. (расчетное число семян – 4752 шт.), а с куста, произрастающего на открытом месте, – 2443 шт. коробочек и 19545 шт. семян. Среднее число коробочек с одного куста C. vulgaris, произрастающего под пологом, составило 209,14 ± 70,2 шт. (расчетное число семян – 1673,14 ± 561,4 шт.), что в 10 раз меньше, чем на открытом месте. С куста, произрастающего на открытом месте, – коробочек 2133,41 ± 768,8 шт. и семян 17067,3 ± 6150,7 шт. соответственно.
Освещенность влияет на формирование числа коробочек, и, как следствие, формируется большое число семян. Так, нами было отмечено, что с увеличением полноты древостоя освещенность под пологом снижалась почти в три раза. W. Beijerinck (1940) показал, что C. vulgaris формирует большее число семенных коробочек, произрастая на хорошо дренированных песчаных почвах с обязательным присутствием мхов. На переувлажненных местообитаниях (болотах) и на вершинах склонов, а также под пологом сухих сосновых лесов, где мхи заменяются лишайниками, число коробочек резко падает.
Объемная влажность субстрата на глубине 10–15 см варьировала от 5,37 до 23,35 % и в среднем составила 11,85 ± 0,6 %.
В верещатниках центральной части ареала под пологом вереска скапливается довольно большое количество опада вереска (Gimingham, 1960), которое, как и на моховом покрове Pleurozium Schreberi, препятствует семенному возобновлению (Sleigh, 1956). Наиболее успешно его семенное возобновление протекает после пожаров, которые освобождают верхние горизонты почвы от подстилки и обеспечивают доступ семян к поверхности почвы (Gimingham, 1960). В ходе многолетних исследований в Англии (Gimingham, 1960) выявлено, что созревшие в огромном количестве семена вереска (в среднем 158000 на 1 м2) падают на подстилку, не заглубляются в почву и проходящий пожар уничтожает их большую часть. Тем не менее оставшихся семян хватает для формирования плотной стены всходов через 12–14 дней после пожара. Для полного восстановления верещатника требуется около 6 и более лет. Успешность восстановления вереска находится в тесной зависимости от температуры горения (Wallace, 1917; Elliott, 1953; Fenton, 1951).
В условиях северной лесостепи Зауралья среднее число семян, находящихся в почве, составляет 104750 ± 48935,9 шт./1 м2, что в 1,5 раза меньше, чем в Шотландии (Gimingham, 1960). По данным европейских авторов, семена вереска, в большом количестве находящиеся в почве (на глубине не более 5 см), могут сохранять свою всхожесть до 100 лет (Mallik, Hobbs, Legg, 1984; Legg, Maltby, Proctor, 1992).
Под пологом леса в сосняках бруснично-вересково-зеленомошных, где C. vulgaris встречается на фоне лишайников рода Cladonia на повышениях микрорельефа, наблюдается мозаичность в обилии его семеношения. В среднем оно составляет 22000 ± 11000 шт./1м2. В понижении на трансекте рельефа, где C. vulgaris растет в синузиях со мхами (Pleurozium Schreberi и Ptilium crista-castrensis Hedw.), число семян возрастает в 8,5 раз – до 187500 ± 23500 шт./1 м2. На открытом местообитании, где вереск также произрастает в окружении мхов, число семян под его сомкнутыми кустами составило в среднем 174500 ± 10500 шт./1 м2, что несколько ниже, чем под пологом леса. Приведенные выше данные подтверждают относительно благоприятные условия для развития C. vulgaris в Западной Сибири как под пологом сосновых лесов, так и на открытых участках в микроассоциациях с мхами.
Влияние субстрата на прорастание семян. В ходе нашего эксперимента по проращиванию семян вереска на различных субстратах (свежая гарь, мох) выявлено, что семена, помещенные в затемненные чашки Петри под моховую подушку, не проросли ни в одном из пяти вариантов (Брянск, Нижний Новогород, Воронеж, Тугулым, Череповец, Курган) (рис. 4, а). В литературе отмечено, что семена вереска светочувствительны и неспособны прорастать в темноте, а также на слое мхов или лишайников (Junttila, 1972; Николаева, 1979; Maltby, 1980; Legg, Stickney, 1989, Maltby, Proctor, 1992; Equihua, Usher, 1993).
Возможно также, что одним из факторов отсутствия прорастания на моховой подушке можно назвать отсутствие микоризы (рис. 4, г). Показано, что заражение грибами происходит из семенной кожуры C. vulgaris через корневой чехлик, а эндосперм и зародыш стерильны. Известно, что семена сосны обыкновенной на свежей гари могут прорастать и без участия грибов, например на свежей гари, где микориза убита, а рН субстрата – 6,5 ± 0,3 (Санников, 1966), при этом процесс их прорастания может замедляться, а сформированные проростки отличаются недоразвитием корневой системы (Rayner, 1913).
В лабораторном эксперименте прорастание семян на свежей гари началось равномерно на 27 сутки, а максимальная всхожесть зафиксирована на 31–34 сутки в Курганской популяции (16 %). Наибольшая энергия прорастания отмечена также у семян из Притоболья (7,3 %). На 40-е сутки эксперимента зарегистрирован всплеск прорастания семян во всех популяциях (рис. 2 и 4, д).
Влияние выжигания мохового покрова. В работах зарубежных авторов отмечено, что незначительное тепловое воздействие несколько ускоряет прорастание семян вереска, но длительное высокотемпературное воздействие (200 °С и более) приводит к гибели семян. Отмечена также взаимосвязь между увеличением температуры пала и снижением доли проросших семян (Kayll, 1968; Equihua, Usher, 1993). Наиболее оптимальные условия, по-видимому, создаются в пределах от 80 до 100–150 °С при длительности прогрева почвы не более 15 минут (Wallace, 1917; Elliott 1953; Fenton, 1951).
В наших экспериментальных посевах семян первые всходы зафиксировали на высечках с открытого местообитания, подверженных экспериментальному выжиганию, уже на 15 сутки (рис. 3), что согласуется с данными европейских авторов (Beijerinck, 1940; Gimingham, 1960). На обожженных почвенных высечках из-под полога древостоя первые всходы появились лишь на 45 сутки. Примерно в это же время появились всходы на высечках, прогретых в сухожарном шкафу (42–50 сутки). Появление первых семядолей в контроле зафиксировано на 21–23 сутки эксперимента.
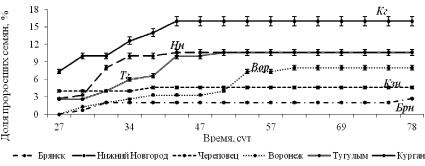
Рис. 2. Динамика прорастания семян C. vulgaris на свежей гари
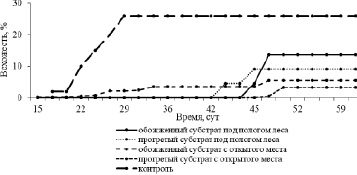
Рис. 3. Динамика прорастания семян C. vulgaris при различных тепловых воздействиях
Максимальное число проросших семян во всех вариантах полевого опыта зафиксировано на 45 сутки, а в контроле при оптимальной влажности субстрата максимум зафиксирован уже на 28 сутки (рис. 3). Пожар, который уничтожил моховой покров и подстилку, в отличие от искусственного прогревания, несколько ускорил прорастание семян, хотя в нашем случае не привел к его значительной стимуляции. Кроме детерминирующей роли влажности субстрата, показанной в полевых экспериментах с семенами P. sylvestris Н.С. Санниковой (1984), в качестве одного из важных факторов следует отметить освещенность, которая, вероятнее всего, оказывает стимулирующее действие на прорастание (Bjorkman, 1945; Leyton, 1955; Jonsson, 1957). Возможно, что отставание семян на 17 суток, произрастающих в открытом грунте, также связано со временем года (Rayner, 1913).

Рис. 4. Иллюстрации проросших семян C. vulgaris на различных субстратах: а – семя под моховой подушкой; б – проросток на моховой подушке; в – проросток на минеральном субстрате (свежая гарь); г – проросток, опутанный гифами грибов, на минеральном субстрате, д – прорастание семени на минеральном субстрате. Примечание: размер линейки на фотографиях равен 1 см
Заключение
Наши исследования в борах северной лесостепи Притоболья показали, что семенная продуктивность реликтовых популяций Calluna vulgaris (L.) Hull в Западной Сибири несколько меньше, чем в европейской части его ареала. При этом под пологом сомкнутого древостоя сосны обыкновенной средняя урожайность семян C. vulgaris на порядок ниже, чем на открытой вырубке, что, вероятно, обусловлено, прежде всего, влиянием корневой и «световой» конкуренции древостоя (Петрова, Мищихина, Черепанова, 2012). Тем не менее уровень семеношения ценопопуляций вереска под пологом леса достаточен для их самовоспроизводства.
Результаты натурных и лабораторных экспериментов свидетельствуют о том, что в сосняках-зеленомошниках успешное прорастание семян и естественное семенное возобновление ценопопуляций вереска возможны лишь после пожаров. Пожары уничтожают слой мха Pleurozium schreberi, который, вероятно, препятствует прорастанию семян вследствие дефицита влажности, отсутствия света под его слоем, куда проникают опадающие семена. Из-за отсутствия микоризы формируется недоразвитая корневая система и замедляется прорастание вереска.
Работа выполнена при поддержке РФФИ (проект № 15-04-03899_а).
Библиографическая ссылка
Петрова И.В., Черепанова О.Е., Кочубей А.А., Санникова Н.С. ОСОБЕННОСТИ СЕМЕНОШЕНИЯ И ПРОРАСТАНИЯ СЕМЯН CALLUNA VULGARIS (L.) HULL В ПРИТОБОЛЬЕ ЗАПАДНОЙ СИБИРИ // Успехи современного естествознания. 2016. № 6. С. 104-109;URL: https://natural-sciences.ru/en/article/view?id=35975 (дата обращения: 14.07.2026).