


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
IMPACT OF ABIOTIC STRESS ON GROWTH OF PLANT

Рост растений в различных геоэкологических условиях позволяет получать информацию о качестве среды произрастания. Армения отличается большим разнообразием геоэкологических зон, интенсивная эксплуатация природных ресурсов в которых в последнее время вызывает определенные риски для окружающей среды. В числе объектов пролонгированного использования природных ресурсов следует отметить горные реки. С другой стороны, присущий данному региону абиотический стресс засухи вносит свои коррективы: Армения занимает 22-е место в списке из 33 стран, которым грозит засуха в течение ближайших 25 лет, отмечается в докладе Всемирного института ресурсов [8].
Засуха способна ограничивать рост растения, вызывая отличия между средним уровнем продуктивности растения в странах с сухим континентальным климатом [5]. При этом снижение доступности воды в почве приводит к уменьшению водного потенциала, который и указывает направление движения воды в почве или в системе растения, а следовательно, и смещение воды в/из растительной клетки.
Известно, что процессы, при помощи которых растительная клетка контролирует рост и формирование, являются результатом тургора и обусловлены напряжением и релаксацией клеточных стенок [4]. Поэтому количественное увеличение воды в клетке приводит к возрастанию объема вакуолей, и, как следствие, клеточный сок давит на цитоплазму, прижимая к клеточной оболочке.
Очевидно, что определение величины тургора прежде всего актуально при экологических исследованиях, так как возможно оценить максимальную способность растения поглощать воду из почвы и удерживать ее. В этом контексте величина тургора может являться одной из геоэкологических характеристик зоны произрастания данного вида растения.
Целью представленной работы является изучение вопросов экологической адаптации, вызванное ограниченным количеством воды в почве для растения кукурузы.
Материалы и методы исследования
В качестве объекта исследования были выбраны инбредные линии кукурузы В73 [3] (контроль – образец 5) и армянской популяции полузубовидной кукурузы. Последние отличаются по основному ареалу произрастания растения на территории Армении. Три образца растения распространены в Лорийском районе Армении вблизи реки Дебет (Шнох – образец 1, Техут – образец 2, Одзун – образец 3) и одно растение – в Армавирском районе Армении вблизи реки Аракс (Ушакерт – образец 4).
Для определения скорости роста растения каждый день измеряли длину его пятого листа. Все образцы растения кукурузы были выращены в камере с контролируемыми климатическими условиями (16 часов день/8 часов ночь, соответственно 25/18 °C; влажность 20 %; фотосинтетически активная радиация 300 mE•м–2•с–1). Контрольные вазоны с семенами поливали каждодневно, поддерживая относительную влажность почвы (ОВП) в вазонах 54 %. При моделировании сильного водного стресса ОВП почвы поддерживали на уровне 34 % (листья увядали в течение дня).
После заметного снижения скорости роста пятого листа во всех образцах определяли общее содержания воды (транспирация). С этой целью срезали всю надземную часть растения, взвешивали и помещали в термостат до полного испарения влаги при температуре +70 °С на 72 часа и затем образцы снова взвешивали. Транспирационный коэффициент определяли по разнице сырого и сухого веса.
Для измерения величины осмотического потенциала воды потенциометрическим методом с помощью прибора PSYPRO (WESCOR Inc.) посредством измерения относительной влажности воздуха над образцом в закрытой камере, который оснащен датчиком С-52 (WESCOR Inc., Logan, UT, США). При равновесии температур относительная влажность воздуха равна значению водного потенциала В начале в измерительную камеру помещали свежесрезанный сегмент листа растения. Затем данный образец был немедленно заморожен помещением в жидкий азот при температуре. Спустя 10 минут образец снова помещался в измерительную камеру и измерялся его осмотический потенциал. Величину тургора в МПа определяли разностью водного и осмотического потенциалов. Все проведенные эксперименты имели 10 биологических и 3–5 технических повторностей. Результаты были обработаны с помощью программы MatLab [9].
Результаты исследования и их обсуждение
В наших исследованиях была поставлена цель выяснить возможные механизмы водопоглощения растениями в различных экологических зонах их произрастания с учетом эффекта засухи.
В начальной стадии экспериментов была определена скорость роста растений по изменению длины его пятого листа. Она измерялась ежедневно в одно и то же время и рассчитывалась как степень удлинения листа (СУЛ) во время стационарного роста растения в течение трех дней после появления.
Полученные результаты представлены на рис. 1, согласно которым армянские образцы отличались по скорости произрастания по сравнению с контролем В73.
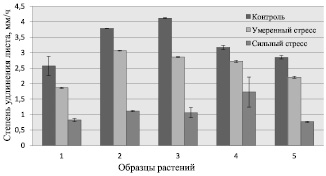
Рис. 1. Скорость роста растений при моделируемой засухе: 1 – образец растения из Шноха; 2 – образец из Техута; 3 – образец из Одзуна; 4 – образец из Ушакерта; 5 – контрольный образец B73
Условно их можно разделить на две группы: быстро (образцы 2 и 3) и медленно (образцы 1 и 4) растущие. Значение СУЛ для растений из первой группы превышало значение по сравнению с контролем (образец 5) в среднем на 38 %, а для второй группы изменение было незначительным. Умеренный стресс вызвал замедление роста у армянских образцов, но, по сравнению с контролем, все же СУЛ было больше в среднем на 15 %. При сильной засухе наблюдалось снижение скорости роста у всех образцов, хотя по сравнению с контрольным растением значение СУЛ уменьшилось в среднем на 4 %. Совершенно иная ситуация наблюдается у образца 4. В обоих случаях моделируемой засухи скорость его роста составила в среднем 37 %. Анализ полученных результатов указывает, что засуха изменяет морфологические показатели у растений, однозначно замедляя их рост.
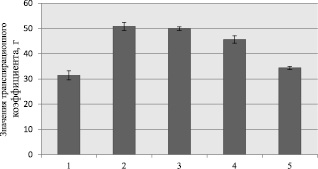
а
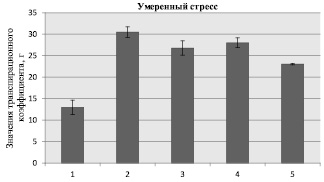
б
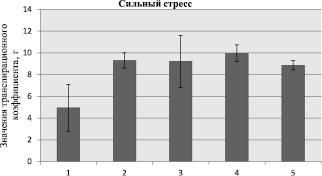
в
Рис. 2. Значения транспирационного коэффициента для образцов кукурузы при моделируемой засухе (обозначение образцов по рис. 1)
Известно, что засуха приводит к дефициту воды, который определяется как дисбаланс между наличием количества воды как в почве, так и ее испарением с поверхности растения [5]. В дальнейших экспериментах было определено значение транспирационного коэффициента, позволяющего оценить количество поглощенной воды надземной частью растения. Полученные результаты представлены на рис. 2, согласно которым, вне зависимости от условий моделируемой засухи, поглощение влаги надземной частью у всех образцов было почти одинаковым и составляло при контрольном условии эксперимента 88 % (рис. 2, а), в случае умеренного стресса 86 % (рис. 2, б), а при жестком стрессе – 76 % (рис. 2, б).
Очевидно, что абиотический стресс – засуха почвы способствует снижению общего содержания влаги в надземной части растений в целом на 39 % при умеренной засухе (рис. 2, б) и на 77 % при сильной засухе (рис. 2, в). После удаления влаги из образцов эти цифры уменьшаются и соответственно составляют 28 и 57 %.
Засуха генерирует физиологические изменения в клетках высших растений, регулируя осмотический потенциал клеток листьев при ограничении доступа воды.
Рост растений обусловленный поглощением воды, регулирует эластичность клеточных стенок [5]. Растения, подверженные дефициту воды, проявляют морфологические изменения, которые являются результатом деформации их клеточной стенки [1]. Вероятно, именно засуха вызывает уменьшение толщины стенок клетки, предотвращая адаптацию гидравлической системы при стрессе [7, 10].
Исходя из вышесказанного былa определенa величина тургора как разница значений водного и осмотического потенциалов свежесрезанного 5-го листа кукурузы при тех же условиях моделируемой засухи. Результаты представлены на рис. 3, согласно которым абиотический стресс вызвал неодинаковую реакцию.
Сильный стресс, связанный с минимальным количеством поглощаемой воды, вызывает упругие деформации и/или примыкающие к упругой деформации. Расширение стенок клетки было больше, когда уровень стресса растений увеличился. Тургор, являясь наиболее важным параметром, контролирует расширение клеток растений. Это может быть связано с разницей в концентрации растворенных в воде веществ между внутренней и внешней сторонами клетки. Действующее на клеточную стенку давление противодействует внутренней силе клетки, и, как следствие этого, клетка находится в состоянии растяжимого напряжения [4, 10].
Тем не менее при росте стенка растительной клетки расширяется за счет тургора, и всякий раз, когда концентрация растворенных веществ в растительной клетке возрастает, давление повышает тургор.
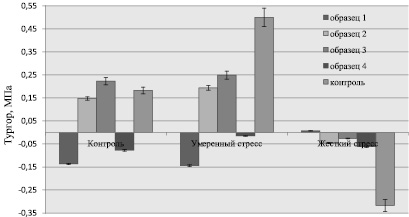
Рис. 3. Сравнение величины тургора при моделируемой засухе в образцах кукурузы: 1 – образец растения из Шноха; 2 – образец из Техута; 3 – образец из Одзуна; 4 – образец из Ушакерта; 5 – контрольный образец B73.
Очевидный факт, что рост растений обусловлен рядом процессов, основой для которых являются поглощение воды и необратимое расширение клеточной стенки. В данном контексте засуха/обезвоживание вне зависимости от природы своего происхождения приводит к стрессу – дефициту воды, который определяется как дисбаланс между наличием воды в почве и испарением. При этом, являясь одним из главных экологических стрессов, засуха способствует замедлению роста растений и продуктивности.
Засуха порождает физиологические изменения в высших растениях, включая потерю тургора, осмотического регулирования и водного потенциала листьев. Отсюда можно заключить, что тургор является той физической силой (давлением), которая и способна вызвать расширение растительной клетки, следовательно, является решающим фактором в регуляции клеточного роста. Во время понижения тургора, вызванного засухой, наблюдается снижение, а в некоторых случаях и прекращение роста растения путем уменьшения растяжимости и расширения его клеток [4]. Механизм этого явления заключается в том, что морфологические изменения в растениях, подвергнутых воздействию дефицита воды, являются результатом изменений клеточных стенок растений. При засухе у саженцев Picea mariana наблюдалось уменьшение толщины клеточной стенки, обусловленное более низким выделением углерода до формирования клеточной стенки, приводящее к торможению адаптационных гидравлических систем при дефиците воды [2]. Хотя следует четко прояснить, что расширение клетки происходит только тогда, когда тургор больше пороговой текучести клеточной стенки. Например, при низких температурах дефицит воды вызывает скручивание листьев, что было связано с фенол-составляющими компонентами клеточной стенки [2]. Хотя механизм адаптации растения допускает потерю определенного количества воды, но при транспирации с поверхности листьев.
Таким образом, величина тургора позволит оценить количество поступившего растворенного в воде вещества в клетку. Очевидно, что транспирация регулируется водой и будет напрямую регулироваться доступностью последней в клетку. Так как растворенные вещества разбавляются в поступающей воде, клетка начинает расширяться и достигать равновесного состояния, при котором укрепляются ее стенки. Данные изменения отображаются и на физиологическом уровне, регулируя скорость роста растения.
Библиографическая ссылка
Сукиасян А.Р., Тадевосян А.В., Симонян Г.С., Пирумян Г.П. ВЛИЯНИЕ АБИОТИЧЕСКОГО СТРЕССА НА РОСТ РАСТЕНИЙ // Успехи современного естествознания. 2016. № 7. С. 168-172;URL: https://natural-sciences.ru/en/article/view?id=36024 (дата обращения: 02.07.2026).