


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
THE QUALITY OF POLLEN OF SPECIES AND REMOTE HYBRIDS IN THE FAMILY GROSSULARIACEAE DC

Фертильность и жизнеспособность пыльцы плодовых культур являются важными показателями, поскольку напрямую связаны с продуктивностью растений. Кроме того, эффективность селекционной работы во многом зависит от качества пыльцы.
По качеству пыльцевых зерен как одному из признаков можно производить отбор наиболее урожайных форм [11].
Известно, что подавляющее большинство современных сортов смородины и крыжовника высокосамоплодны и имеют нормально развитую пыльцу с высокой оплодотворяющей способностью [1; 2; 4; 6; 10].
Вопросы биологии пыльцы отдаленных гибридов в семействе Grossulariaceae DC изучены недостаточно полно. По сведениям Е. Кип [4] аллотетраплоиды отличаются по фертильности в соответствии со степенью близости родительских видов. У тетраплоидов от фертильных внутрисекционных гибридов фертильность сильно снижена, но тетраплоидные формы стерильных межсекционных гибридов обычно обладают хорошей фертильностью. Исследованиями И.Э. Бученкова [1]; И.Э. Бученкова, О.С. Рышкель и др. [2] показано, что при переводе диплоидных сортов на тетраплоидный уровень фертильность снижается в среднем у черной смородины в 2,3 раза; у крыжовника – в 1,36 раза. Е.В. Ульяновской, В.В. Ковалевой и др. [9] приводятся данные о том, что пыльца триплоидных сортов прорастает либо единичными пыльцевыми трубками, либо не прорастает совсем.
Целью настоящего исследования явилась оценка качества пыльцы, а также изучение особенностей формирования пыльцевых зерен у отдаленных гибридов смородины и крыжовника и их исходных диплоидных форм.
Материалы и методы исследования
Объектами исследования служили виды рода Ribes L.: R. nigrum subsp. europaeum Jancz., R. aureum Purch, R. spicatum E. Robson; виды рода Grossularia Mill: G. reclinata (L.) Mill., G. divaricata (Dougl.) Cov. et Britt., G. nivea Spach; аллотетраплоидные смородинно-крыжовниковые гибриды: Josta, Kroma, B 1323/3 и 3231 и отдаленные гибриды: R. hudsonianumR. dikuscha; 046 Petroc 69; 048 Petjancz 33.
Гибриды, включенные в исследование, имеют различное генетическое и эколого-географическое происхождение. Аллотетраплоид Josta был создан в 1970-х гг. в институте им. Макса Планка (ФРГ). Исходными формами служили (R. nigrum×G. reclinata)×(R. nigrum×G. divaricata). Гибрид Kroma был выведен в Швеции, на сельскохозяйственной станции в Альнарпе от скрещивания (R. nigrum×Grossularia)×(R. nigrum×G. nivea). В настоящее время они получили статус сортов и успешно культивируются в странах Западной Европы [7].
Смородинно-крыжовниковые гибриды В 1323/3 и 3231 были получены на Ист-Моллингской станции (Великобритания). Генетическое происхождение их не установлено.
Отдаленные межвидовые и межподвидовые гибриды R. hudsonianum×R. dikuscha; 046 Petroc 69 и 048 Petjancz 33 были созданы А.И. Рилишкисом (Литва, Каунас). Исходными формами при получении 046 Petroc 69 служили R. petraeum×R. procumbens; 048 Petjancz 33 – R. petraeum×R. janczewskii. Родительскими формами триплоида Длиннокистная ЦГЛ (2n = 24) являются: Кызырган (R. altissimum)×Восьмая Дэвисона (R. nigrum subsp. europaeum). Он был получен во Всероссийском НИИ генетики и селекции плодовых растений (ВНИИГиСПР им. И.В. Мичурина, бывш. ЦГЛ им. И.В. Мичурина).
Пыльцу для определения ее фертильности и жизнеспособности собирали в период массового цветения на коллекции черной смородины НПБ «Пушкинские и Павловские лаборатории ВИР» и в коллекции ботанического сада БИН РАН.
Микроскопические исследования проводили в лаборатории палинологии БИН РАН с помощью светооптического микроскопа Микмед-6 при увеличениях 20×10 и 40×10. Фертильность пыльцевых зерен изучали с помощью традиционного ацетокарминового метода. Для определения процента фертильных и стерильных пыльцевых зерен проводили подсчет количества зерен не менее, чем в 10 полях зрения. Жизнеспособность пыльцы оценивали путем проращивания на агаризированной среде, содержащей 1 % агар-агар, 0,01 % раствор борной кислоты и 0,5 мол/л раствор сахарозы (твердая среда) и на 15 % растворе сахарозы (жидкая среда). Жизнеспособной считали пыльцу, размер пыльцевой трубки которой превышал величину пыльцевого зерна.
Для изучения особенностей формирования пыльцы просматривали стадии ее развития путем приготовления давленых препаратов каждые 2–4 дня от начала вегетации до момента выдвижения кистей.
Результаты исследования и их обсуждение
Исследования, проведенные нами ранее, показали, что пыльцевые зерна смородин и крыжовников среднего размера (25–40 мкм в диаметре). Они являются весьма специфичными, обладают рядом признаков, характерных для семейства крыжовниковых в целом, и довольно четко отличаются друг от друга и от пыльцы других семейств в спорово-пыльцевых спектрах [12]. Смородинно-крыжовниковые гибриды имеют зерна с переходными апертурными типами, содержащими и борозды, и поры. Кроме того, у тетраплоидных гибридов выявлен больший разброс размеров пыльцевых зерен [3].
В исследование были включены виды с глобально расположенными порово-оровыми пыльцевыми зернами – правильными (R. nigrum subsp. europaeum, R. aureum) и отклоняющимися (R. spicatum) вариантами, и видов с меридионально расположенными бороздно-оровыми зернами – также с правильными (G. reclinata, G. nivea) и отклоняющимися (G. divaricata) вариантами, а также гибридов с правильными (048 Petjancz 33), неправильными (R. hudsonianum×R. dikuscha; 046 Petroc 69) глобально порово-оровыми и переходными (смородинно-крыжовниковые гибриды: Josta, Kroma, B 1323/3 и 3231) апертурными типами.
Исследованиями Н.П. Чувашиной [8] была выявлена корреляция между фертильностью и образованием тетрад и/или полиад в ходе микроспорогенеза.
Для онтогенетического исследования были выбраны тетраплоидные смородинно-крыжовниковые гибриды Kroma и Josta и их известные родительские виды (R. nigrum, G. reclinata, G. divaricata, G. nivea), а также триплоид Длиннокистная ЦГЛ. Изучение особенностей формирования пыльцы этих образцов показало, что во время вегетации растений, от появления зеленого конуса и до момента выдвижения кистей происходит микроспорогенез (дифференциация спорогенной ткани, мейоз, образование и распад тетрад и/или полиад) и образование отдельных пыльцевых зерен. Стадии раз вития пыльцевых зерен у изученных образцов представлены в табл. 1.
Таблица 1
Стадии микроспорогенеза у представителей семейства Grossulariaceae (2016 г.)
|
Название образца |
Начало вегетации |
Даты |
|||||||
|
07.04 |
11.04 |
14.04 |
16.04 |
18.04 |
20.04 |
22.04 |
26.04 |
||
|
Grossularia nivea |
30.03 |
Сп. ткань |
Сп. ткань |
Тетрады |
Тетрады и отдельные п.з. |
Отдельные п.з. |
|||
|
G. divaricata |
1.04 |
– « – |
– « – |
Тяжи формирующихся тетрад и тетрады |
Тяжи формирующихся тетрад, тетрады, редкие отдельные п.з. |
Отдельные п.з. |
|||
|
G. reclinata |
31.03 |
– « – |
– « – |
– « – |
Тяжи формирующихся тетрад, редкие тетрады |
Тяжи формиру-ющихся тетрад, тетрады, редкие отдельные п.з. |
Отдельные п.з. |
||
|
Kroma |
31.03 |
– « – |
– « – |
Неоднородная спорогенная ткань, тяжи формирующихся тетрад |
Тетрады, полиады |
Тетрады, полиады |
Отдельные п.з |
||
|
Josta |
31.03 |
– « – |
– « – |
Более интенсивно окрашенные тяжи |
Тяжи формиру-ющихся тетрад, редкие тетрады и полиады |
Тяжи формиру-ющихся тетрад, редкие тетрады и полиады |
Отдельные п.з., присутствуют ед. тетрады и полиады |
Отдельные п.з. |
|
|
Ribes nigrum |
5.04 |
– « – |
– « – |
Спорогенная ткань, тяжи |
Спорогенная ткань, тяжи, тетрады |
Тяжи формиру-ющихся тетрад, тетрады |
Тетрады и отдельные п.з. |
Отдельные п.з. и тетрады |
|
|
Дл/кистная ЦГЛ |
30.03 |
– «– |
– «– |
Неоднородная спорогенная ткань, тяжи формирующихся тетрад |
Тяжи формирующихся тетрад |
Тяжи формиру-ющихся тетрад, отдельные мелкие п.з. |
Тетрады, отдельные мелкие и крупные п.з., присутствуют тяжи |
Отдельные п.з., присутствуют тетрады и тяжи |
Отдельные п.з., присутствуют тетрады и тяжи |
Стадия тетрад у изучаемых образцов выявлена в период с 14 по 26 апреля. Раньше всех эту стадию (спустя 10–13 дней от начала вегетации), а также стадию образования отдельных пыльцевых зерен (через 17 дней от момента выдвижения зеленого конуса) проходят крыжовники, в первую очередь G. nivea и G. divaricata (табл. 1). У смородинно-крыжовниковых гибридов формирование отдельных пыльцевых зерен происходит cпустя 3 недели от начала вегетации с небольшой разницей в 2 дня (табл. 1), причем у сорта Josta на протяжении всего периода онтогенеза можно было наблюдать одновременно все его стадии вплоть до момента образования распавшихся отдельных пыльцевых зерен (22.IY). У смородин – европейского подвида R. nigrum и триплоида Длиннокистная ЦГЛ стадии онтогенеза пыльцы проходят позже, чем у крыжовников и смородинно-крыжовниковых гибридов.
Следует отметить, что почти у всех изученных образцов в поле зрения микроскопа можно было наблюдать одновременно наличие разных стадий формирования пыльцевых зерен (например, стадии тетрад, распадающихся тетрад и отдельных пыльцевых зерен, рис. 1, г).
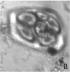
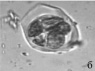
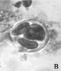
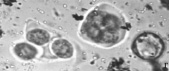
Риc. 1. Микроспорогенез у изучаемых образцов: а) полиада (Josta), б) полиада (Kroma), в) тетрада (G. nivea), г) тетрада, распадающаяся тетрада и отдельное пыльцевое зерно (R. nigrum)×400
В особенности в течение довольно длительного времени это было характерно для гибридов. Среди изученных нами образцов образование полиад выявлено только у гибридов. При воздействии ацетокармином обнаружено, что многие полиады окрашиваются полностью, а в некоторых тетрадах отдельные формирующиеся микроспоры остаются неокрашенными. Пыльцевые зерна в этот период остаются незрелыми, их формирование продолжается до начала цветения.
В проведенное нами исследование фертильности пыльцы помимо перечисленных образцов были включены смородинно-крыжовниковые гибриды В1323/3 и 3231, отдаленные межвидовые гибриды R. hudsonianum×R. dikuscha; 046 Petroc 69, 048 Petjancz 33 и виды с правильным (R. aureum) и неправильным расположением апертур (R. spicatum). Результаты представлены в табл. 2.
Таблица 2
Количественное содержание фертильных и стерильных пыльцевых зерен в зрелых пыльниках видов и отдаленных гибридов семейства Grossulariaceae
|
Название образца |
Количество просмотренных пыльцевых зерен, шт. |
Фертильные п.з. |
Аномальные п.з. |
Особенности |
||
|
Общее количество, шт. |
% от общего количества |
Общее количество, шт. |
% от общего количества |
|||
|
G. divaricata |
195 |
190 |
97,4 |
5 |
2,6 |
|
|
G. nivea |
214 |
200 |
93,5 |
14 |
6,5 |
|
|
R. nigrum subsp. europaeum |
282 |
260 |
92,2 |
22 |
7,8 |
|
|
048 Petjancz 33 |
591 |
545 |
92,2 |
46 |
7,8 |
|
|
R. aureum |
413 |
373 |
90,3 |
40 |
9,7 |
Неокрашенные – главным образом мелкие |
|
R. hudsonianum×R.dikuscha |
271 |
236 |
87,1 |
35 |
12,9 |
Неокрашенные – мелкие и средние |
|
В 1323/3 |
382 |
317 |
82,9 |
65 |
17,1 |
|
|
Kroma |
515 |
396 |
76.9 |
119 |
23,1 |
Неокрашенные – мелкие и средние |
|
G. reclinata |
398 |
283 |
71,1 |
115 |
28,9 |
|
|
Josta |
529 |
360 |
68,1 |
169 |
31,9 |
|
|
3231 |
589 |
386 |
65,5 |
203 |
34,5 |
|
|
R. spicatum |
212 |
118 |
55,7 |
94 |
44,3 |
Неокрашенные – мелкие, сгруппированы по 3–8 зерен |
|
046 Petroc 69 |
641 |
323 |
50,4 |
318 |
49,6 |
|
|
Длиннокистная ЦГЛ |
897 |
49 |
5,5 |
848 |
94,5 |
|
Определение фертильности пыльцы, проведенное нами, показало, что доля морфологически зрелой (фертильной) пыльцы от общего количества просмотренных пыльцевых зерен составила в зависимости от образца 5,5–97,4 % (табл. 2). Наиболее высокий уровень фертильности (90,3–97,4 %) наблюдался главным образом у образцов с правильным расположением апертур – G. nivea, R. nigrum subsp. europaeum, R. aureum, 048 Petjancz 33, а также у G. divaricata, апертуры пыльцевых зерен которого расположены неравномерно. Достаточно высокая степень морфологически зрелой пыльцы (82,9–87,1 %) была характерна для гибридов с большим количеством пыльцевых зерен, отклоняющихся от типичных с правильными апертурами – R. hudsonianum×R. dikuscha и смородинно-крыжовникового гибрида В 1323/3. Остальные смородинно-крыжовниковые гибриды, наряду с G. reclinata характеризовались средним уровнем фертильности (65,5–76,9 %). У образцов с неправильным расположением апертур (046 Petroc 69, R. spicatum) показатель фертильности был относительно невысоким (50,4–55,7 % соответственно). Самый низкий уровень фертильности был свойственен триплоиду Длиннокистная ЦГЛ. Характерные состояния пыльцы в полях зрения микроскопа от высокого уровня фертильности к низкому показаны на рис. 2.
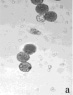
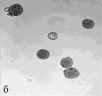
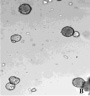
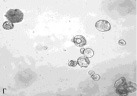
Рис. 2. Пыльцевые зерна, окрашенные ацетокармином: а) 048 Petjancz 33, б) Ribes hudsonianum×R .dikuscha, в) 046 Petroc 69, г) Длиннокистная ЦГЛ, х 200
В основном неокрашенными оставались мелкие зерна, иногда только мелкие (10–15 мкм в диаметре). Стерильные пыльцевые зерна под микроскопом выглядели сморщенными, деформированными, мелкими по величине (R. aureum); мелкими и средними (R. hudsonianum×R. dikuscha); мелкими, сгруппированными по 3–8 зерен (R. spicatum).
Важными характеристиками пыльцы являются длина пыльцевой трубки и скорость ее роста, поскольку обеспечивают тот или иной уровень конкурентоспособности при прорастании на рыльце пестика [5].
Исследования показали, что жизнеспособность пыльцы изученных гибридов была существенно ниже уровня фертильности и не превышала 30 %. Выявлено, что прорастание пыльцы у образца 048 Petjancz 33 происходит уже через 4 часа, причем у проросших зерен формируются длинные пыльцевые трубки (рис. 3, а). В отличие от указанного образца пыльцевые трубки других отдаленных гибридов (R. hudsonianum×R. dikuscha и 046 Petroc 69) были короче. Спустя сутки наблюдалась примерно та же картина, что и ранее (через 4 часа) – количество проросших пыльцевых зерен не увеличилось. Выявлено, что на жидкой среде прорастание пыльцевых зерен происходит более интенсивно у всех изученных образцов. Пыльца тетраплоидных смородинно-крыжовниковых гибридов прорастает. одновременно из двух апертур (проростковых пор), тем самым образуя две пыльцевые трубки (рис. 3, б). У триплоида прорастающих пыльцевых зерен не обнаружено.
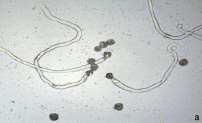
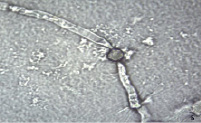
Рис. 3. Прорастание пыльцевых зерен на искусственной питательной среде: а) 048 Petjancz 33 х 200, б) Kroma, x 400
Заключение
Изученные образцы – виды и отдаленные гибриды, в том числе тетраплоидные, за исключением триплоидного, обладают высоким уровнем фертильности, что свидетельствует о возможности их использования в селекции. У изученных смородинно-крыжовниковых гибридов процент фертильных пыльцевых зерен ниже в 1,2–1,4 раза, чем у их родительских форм (R. nigrum subsp. europaeum, G. divaricata, G. nivea). Не выявлено существенных различий по уровню фертильности у образцов с разным типом апертур.
У гибридов в процессе микроспорогенеза происходит образование как тетрад, так и полиад. Корреляция между фертильностью и образованием полиад слабая.
Жизнеспособность пыльцы изученных образцов ниже уровня их фертильности.
При прорастании тетраплоидных смородинно-крыжовниковых гибридов вначале одновременно образуются две пыльцевые трубки (из двух апертур – проростковых пор), что отличает их от других изученных отдаленных гибридов, у которых сразу формируется одна пыльцевая трубка.
Исследование выполнено при финансовой поддержке Российского фонда фундаментальных исследований, проект № 15-04-06386_а.
Библиографическая ссылка
Гаврилова О.А., Тихонова О.А. КАЧЕСТВО ПЫЛЬЦЫ ВИДОВ И ОТДАЛЕННЫХ ГИБРИДОВ В СЕМЕЙСТВЕ GROSSULARIACEAE DC // Успехи современного естествознания. 2016. № 12-1. С. 68-73;URL: https://natural-sciences.ru/en/article/view?id=36263 (дата обращения: 24.06.2026).