


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
PHOTOSYNTHETIC ACTIVITY OF PLANTS IN CROPS AND DYNAMICS OF GROWTH PROCESSES IN THE APPLICATION OF BIOLOGICAL PRODUCTS

Взаимосвязь растений в агроценозе носит непостоянный характер, зависящий от многих факторов. Главной задачей для получения высоких урожаев является создание такого посева, в котором бы максимально раскрывались потенциальные возможности фотосинтетической деятельности растений в агроценозе. Этого можно добиться при создании благоприятных условий для роста и развития растений.
Ряд авторов [3, 14] рассматривает рост растений как процесс дифференцирования организма за счет образования новых и увеличения старых элементов его структуры (молекул, клеток, тканей и органов), оказывающий решающее влияние на распределение, перераспределение и использование образовавшихся при фотосинтезе и метаболизме органических веществ, а также поглощение минеральных солей и воды, идущих на образование поверхности органов и тканей, их регенерацию и на запасные отложения. Рост растения суммирует в себе и венчает многие другие процессы жизнедеятельности растения, выражая в известной мере баланс процессов синтеза и распада веществ в организме при его взаимодействии с условиями внешней среды.
Рост растения и его биологическая продуктивность – результат, прежде всего, фотосинтетической деятельности, в ходе которой образуется до 95 % органических соединений. Поэтому рост растения, формативный, органообразовательный и рост как увеличение сухой биомассы начинается, главным образом, вслед за формированием фотосинтетической системы листа и осуществления процесса фотосинтеза. Лист как орган фотосинтеза является центром образования первичных продуктов, их метаболизации и эвакуации в органы запаса [2, 15].
Целью данной работы являлось выявление особенностей формирования фотосинтетической деятельности растений в посевах и динамика ростовых процессов при применении биологических препаратов и минеральных удобрений.
Материалы и методы исследования
Организация полевых опытов, проведение наблюдений и лабораторных анализов осуществлялось по общепринятым методикам, изложенным в следующих источниках: «Методика опытного дела» [1], «Фотосинтетическая деятельность растений в посевах» [11], «Оценка эффективности микробных препаратов в земледелии» [12].
Проводили следующие учеты и анализы:
– фенологические наблюдения согласно методике государственного сортоиспытания [13];
– площадь листовой поверхности определялась методом «высечки» на 10 растениях с делянки на двух несмежных повторностях по фазам развития растений по формуле
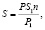
где S – общая площадь листьев одного растения, см2; S1 – площадь одной высечки, см2; P – общая масса листьев одного растения, г; P1 – масса высечек, г; n – число высечек. Зная густоту стояния растений на 1 га рассчитывалась площадь листьев на 1 га;
– содержание сухого вещества определялось отбором средней пробы измельченных растений. Из каждого образца отбирались навески по 50 г в двукратной повторности, фиксировались и высушивались в сушильном шкафу при температуре 105 °С до постоянной массы. Содержание сухого вещества определялось по формуле
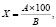
где А – масса навески после высушивания, г; В – масса навески до высушивания, г;
– чистая продуктивность фотосинтеза вычислялась по формуле Кидда, Веста и Бриггса:
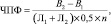
где ЧПФ – чистая продуктивность фотосинтеза; В1 и В2 – сухая биомасса пробы урожая в начале и конце учетного периода, г; В2 – В1 – прирост сухой массы за учетный период n дней, г; (Л1 + Л2)/0,5 – средняя работавшая площадь листьев за этот промежуток времени, м2; n – число дней.
Минеральные удобрения аммиачная селитра, двойной гранулированный суперфосфат и хлористый калий вносили под предпосевную культивацию. Обработка семян бактериальными препаратами и препаратом ЖУСС-2 (жидкий удобрительно-стимулирующий состав) проводили за 1–2 дня до посева. Повторность опыта 4-кратная. Учетная площадь делянок 42 м2 (2,1*20).
Результаты исследования и их обсуждение
Формирование фотосинтетического аппарата представляет сложный процесс. В ранние фазы роста и развития преобладают процессы новообразования и роста листьев, а в более поздние – процессы отмирания, связанные с усиленной транспортировкой пластических веществ в репродуктивные органы [9]. Накопление и запасание энергии в процессе фотосинтеза сопровождается накоплением биомассы, служащей структурным и энергетическим материалом, обеспечивающим существование растений [15].
Максимальные урожаи могут быть сформированы посевами с оптимальной площадью листьев, причем важно, чтобы она быстро нарастала до максимальной величины и долго удерживалась на достигнутом уровне без резкого снижения к концу вегетации, максимально поглощая солнечную радиацию [10]. У культур, потенциально способных к формированию очень большой листовой поверхности, к которым относятся, в частности, злаки [8], степень ее фактического развития, а тем самым и факторы фотосинтетической деятельности, лимитирующие урожайность, решающим образом зависят от условий выращивания. Если ресурсы влаги и питания недостаточны, то основным фактором, лимитирующим урожайность растений, является недостаточное развитие листовой поверхности, а на бедных почвах – и низкая производительность ее работы.
Основными показателями фотосинтетической деятельности растений являются: площадь листьев, фотосинтетический потенциал, чистая продуктивность фотосинтеза и коэффициент использования фотосинтетически активной радиации. Величина площади листьев слагается из площади листьев отдельных растений и в различной степени зависит от периода вегетации, погодных условий выращивания, сорта, густоты растений, обеспеченности их питательными веществами и т.д.
Исследования [4, 5] указывают, что более продуктивным может быть то растение, которое отличается менее интенсивным фотосинтезом, но больший процент ассимилятов использует на образование листьев и формирует большую ассимиляционную поверхность. Поэтому очень важно агротехническими мероприятиями в наиболее короткие сроки добиваться оптимальной площади листьев растений.
Имеется немало указаний в литературе, что с увеличением ассимиляционной площади листьев увеличивается урожай. Однако положительная связь этих двух процессов имеет предел, при котором большая листовая поверхность из-за взаимного затенения снижает интенсивность фотосинтеза, в результате чего увеличивается непродуктивная часть урожая и уменьшается продуктивная.
Наши исследования показывают, что в начале развития растений яровой пшеницы от появления всходов до фазы кущения только начинают проявляться различия вариантов по площади листьев. Площадь листьев в фазу кущения изменяется в пределах 7,5–8,9 тыс. м2/га, а на фоне минеральных удобрений 9,1–11,4 тыс. м2/га (табл. 1). При инокуляции семян наблюдается тенденция к увеличению ассимиляционной поверхности листьев, особенно это заметно при применении бактофосфин + ЖУСС-2. По-видимому, в данный период действие биопрепаратов только начинает проявляться [6].
Таблица 1
Влияние биопрепаратов на ассимиляционную поверхность листьев, тыс. м2/га (среднее за 3 года)
|
Варианты |
Фазы развития |
||
|
кущение |
трубкование |
колошение |
|
|
1. Контроль |
7,5 |
26,2 |
36,6 |
|
2. Азотовит |
8,6 |
28,7 |
39,5 |
|
3. Бактофосфин |
8,1 |
29,5 |
40,7 |
|
4. ЖУСС-2 |
8,7 |
30,7 |
41,5 |
|
5. Азотовит + бактофосфин |
8,4 |
29,7 |
40,9 |
|
6. Азотовит + ЖУСС-2 |
8,5 |
30,7 |
40,7 |
|
7. Бактофосфин + ЖУСС-2 |
8,9 |
31,6 |
42,9 |
|
8. Азотовит + бактофосфин + ЖУСС-2 |
8,1 |
28,9 |
40,5 |
|
9. N30P30K30 (Фон) |
9,1 |
28,9 |
40,4 |
|
10. Фон + азотовит |
9,8 |
31,2 |
42,6 |
|
11. Фон + бактофосфин |
9,8 |
31,1 |
42,4 |
|
12. Фон + ЖУСС-2 |
10,6 |
33,3 |
44,3 |
|
13. Фон + азотовит + бактофосфин |
9,8 |
32,2 |
43,8 |
|
14. Фон + азотовит + ЖУСС-2 |
11,2 |
34,1 |
45,7 |
|
15. Фон + бактофосфин + ЖУСС-2 |
11,4 |
34,1 |
46,2 |
|
16. Фон + азотовит + бактофосфин + ЖУСС-2 |
10,1 |
32,2 |
44,2 |
В фазе трубкования наблюдался более интенсивный рост и существенное увеличение ассимиляционной поверхности листьев по всем вариантам и варьировалось от 26,2 до 31,6 тыс. м2/га, а на фоне удобрений – от 28,9 до 34,1 тыс. м2/га. Более интенсивное нарастание листовой поверхности наблюдалось при инокуляции семян биопрепаратами. Так, увеличение площади листьев на этих вариантах относительно контроля составляло 9,5–20,6 % (фон N0P0K0) и 7,6–18,0 % (фон N30P30K30). В фазе колошения площадь листовой поверхности находилась в пределах 36,6–42,9 тыс. м2/га, а на удобренном фоне 40,4–46,2 тыс. м2/га. При этом на всех исследуемых вариантах этот показатель был выше по сравнению с контролем на 7,9–17,2 %, а на фоне минеральных удобрений – 5,0–14,4 %. Нами отмечено, что интенсивность нарастания листовой поверхности в течение всей вегетации достигала максимальных значений при инокуляции семян бактофосфином в смеси с ЖУСС-2 на фоне минеральных удобрений.
Показатель площади листьев не всегда дает полную характеристику фотосинтетической деятельности посева, поскольку исключительно важно время, когда сформировалась максимальная площадь листьев и сколько дней она работала на накопление урожая.
Важнейшим показателем характеристики продолжительности фотосинтетической работы посева за всю вегетацию или определенный период является показатель – фотосинтетический потенциал посева (ФПП), характеризующий собой сумму ежесуточных показателей площади листьев на гектар посева и выражается в м2·день/га [7, 8].
Наши исследования показали, что при применении биопрепаратов интенсивность нарастания ФПП в течение вегетации яровой пшеницы возрастает за счет увеличения листовой поверхности. Так, в фазу кущения ФПП при значениях на контроле – 0,083 млн м2·дней/га по вариантам варьировался от 0,090 до 0,099 млн м2·дней/га (табл. 2). Такая же закономерность прослеживалась и в дальнейшем.
Таблица 2
Влияние биопрепаратов на фотосинтетический потенциал посевов, млн·м2·дней/га (среднее за 3 года)
|
Варианты |
Фазы развития |
||
|
кущение |
трубкование |
колошение |
|
|
1. Контроль |
0,083 |
0,196 |
0,431 |
|
2. Азотовит |
0,095 |
0,216 |
0,469 |
|
3. Бактофосфин |
0,090 |
0,219 |
0,483 |
|
4. ЖУСС-2 |
0,097 |
0,229 |
0,496 |
|
5. Азотовит + бактофосфин |
0,093 |
0,222 |
0,487 |
|
6. Азотовит + ЖУСС-2 |
0,094 |
0,228 |
0,490 |
|
7. Бактофосфин + ЖУСС-2 |
0,099 |
0,236 |
0,511 |
|
8. Азотовит + бактофосфин + ЖУСС-2 |
0,090 |
0,216 |
0,476 |
|
9. N30P30K30 (Фон) |
0,101 |
0,221 |
0,476 |
|
10. Фон + азотовит |
0,107 |
0,238 |
0,509 |
|
11. Фон + бактофосфин |
0,109 |
0,238 |
0,506 |
|
12. Фон + ЖУСС-2 |
0,117 |
0,255 |
0,534 |
|
13. Фон + азотовит + бактофосфин |
0,109 |
0,245 |
0,522 |
|
14. Фон + азотовит + ЖУСС-2 |
0,124 |
0,263 |
0,548 |
|
15. Фон + бактофосфин + ЖУСС-2 |
0,127 |
0,265 |
0,552 |
|
16. Фон + азотовит + бактофосфин + ЖУСС-2 |
0,112 |
0,246 |
0,525 |
В фазе трубкования на контроле ФПП составил 0,196 млн м2·дней/га; на исследуемых вариантах находился в пределах от 0,216 до 0,236 млн м2·дней/га. В фазу колошения фотосинтетический потенциал посевов повышается на 8,8–18,6 % относительно контроля. Внесение минеральных удобрений оказало существенное влияние на интенсивность нарастания ФПП, но закономерность при этом сохранилась.
Рост и фотосинтез растений составляют основу единого продукционного процесса, и основным показателем фотосинтетической продуктивности растений является накопление ими сухой массы в пересчете на единицу сплошной листовой поверхности за определенный период [2, 7].
Ведущая роль в формировании урожайности принадлежит чистой продуктивности фотосинтеза (ЧПФ), которая характеризует активность работы ассимиляционной поверхности листьев в течение вегетации. В связи с этим продуктивность фотосинтеза широко изучается в различных почвенно-климатических условиях для выяснения факторов, способствующих повышению продуктивности культурных растений и лимитирующих ее продуктивность.
Результаты наших исследований показывают, что в среднем за годы исследований в течение вегетации яровой пшеницы чистая продуктивность фотосинтеза шла по восходящей кривой и колебалась от 2,3 до 6,9 г/м2·сутки в зависимости от минеральных удобрений, биопрепаратов и препарата ЖУСС-2. В фазу кущения продуктивность фотосинтеза колебалась от 2,3 до 2,9 г/м2·сутки, а на фоне минеральных удобрений – от 2,7 до 3,3 г/м2·сутки. Инокуляция семян способствует увеличению ЧПФ на 0,1–0,7 г/м2·сутки.
Рядом авторов установлено, что по мере увеличения в посевах площади листьев ЧПФ падает. Однако имеются и противоположные мнения. Результаты наших исследований показывают, что увеличение площади листьев не приводило к резкому снижению продуктивности фотосинтеза. Это можно объяснить тем, что максимальная площадь листьев в фазу колошения составляла 46,2 тыс. м2/га, что является оптимальной. А.А. Ничипорович [7, 8] считает предельной площадь листьев пшеницы 40–50 тыс. м2/га, если площадь выше этой величины, то листья затеняются и снижается интенсивность фотосинтеза. В фазу трубкования продуктивность фотосинтеза повышается от инокуляции на 2,2–13,3 % (табл. 3).
Таблица 3
Влияние биопрепаратов на чистую продуктивность фотосинтеза, г/м2·сутки (среднее за 3 года)
|
Варианты |
Фазы развития |
||
|
кущение |
трубкование |
колошение |
|
|
1. Контроль |
2,3 |
4,5 |
5,8 |
|
2. Азотовит |
2,5 |
4,6 |
6,2 |
|
3. Бактофосфин |
2,5 |
4,8 |
6,2 |
|
4. ЖУСС-2 |
2,8 |
5,0 |
6,3 |
|
5. Азотовит + бактофосфин |
2,6 |
4,7 |
6,0 |
|
6. Азотовит + ЖУСС-2 |
2,9 |
5,1 |
6,5 |
|
7. Бактофосфин + ЖУСС-2 |
2,9 |
5,1 |
6,5 |
|
8. Азотовит + бактофосфин + ЖУСС-2 |
2,6 |
4,7 |
6,2 |
|
9. N30P30K30 (Фон) |
2,7 |
4,8 |
6,3 |
|
10. Фон + азотовит |
2,8 |
4,9 |
6,5 |
|
11. Фон + бактофосфин |
2,9 |
5,1 |
6,6 |
|
12. Фон + ЖУСС-2 |
3,0 |
5,2 |
6,6 |
|
13. Фон + азотовит + бактофосфин |
3,1 |
5,1 |
6,6 |
|
14. Фон + азотовит + ЖУСС-2 |
3,1 |
5,3 |
6,8 |
|
15. Фон + бактофосфин + ЖУСС-2 |
3,3 |
5,4 |
6,9 |
|
16. Фон + азотовит + бактофосфин + ЖУСС-2 |
3,0 |
5,1 |
6,6 |
В течение вегетации показатели ЧПФ изменялись на фоне минеральных удобрений. Максимальные величины ЧПФ наблюдались в фазе колошения, т.е. в период наибольшего развития листовой поверхности растений яровой пшеницы. Наибольшие величины продуктивности фотосинтеза были на вариантах «азотовит + ЖУСС-2» и «бактофосфин + ЖУСС-2» и составили 6,5 г/м2·сутки, на фоне минеральных удобрений соответственно 6,8 и 6,9 г/м2·сутки.
Заключение
Таким образом, исследования показали, что на характер формирования листовой поверхности яровой пшеницы минеральные удобрения и биопрепараты оказали непосредственное воздействие. Инокуляция семян биопрепаратами увеличивает данный показатель фотосинтетической деятельности, вероятно, за счет стимулирующего действия микроорганизмов, входящих в состав биопрепаратов, которая начинает проявляться с фазы кущения. Как при применении минеральных удобрений, так и без них, наибольшую площадь листьев имели растения на варианте «бактофосфин + ЖУСС». Инокуляция семян способствует увеличению фотосинтетического потенциала посевов в течение вегетации за счет увеличения листовой поверхности. Инокуляция на фоне минеральных удобрений дает возможность формирования более высокого фотосинтетического потенциала посевов яровой пшеницы. При инокуляции семян яровой пшеницы биопрепаратами и препаратом ЖУСС-2 проявляется тенденция к повышению чистой продуктивности фотосинтеза как на неудобренном фоне, так и на фоне минеральных удобрений.
Библиографическая ссылка
Никитин С.Н. ФОТОСИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ РАСТЕНИЙ В ПОСЕВАХ И ДИНАМИКА РОСТОВЫХ ПРОЦЕССОВ ПРИ ПРИМЕНЕНИИ БИОЛОГИЧЕСКИХ ПРЕПАРАТОВ // Успехи современного естествознания. 2017. № 1. С. 33-38;URL: https://natural-sciences.ru/en/article/view?id=36338 (дата обращения: 17.07.2026).