


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
DEVELOPMENT OF MURAVYOV-AMURSKY PENINSULA LANDSCAPES DURING MIDDLE-LATE HOLOCENE BASED ON RESEARCH OF COAST MURAVYINAYA BAY SEDIMENTS DATA

Полуостров Муравьева-Амурского является одним из наиболее освоенных участков Приморского края и целиком входит в границы агломерации «Большой Владивосток». Еще 150 лет назад, до начала интенсивного освоения территории, в окрестностях Владивостока «росли смешанные леса», в которых «чаще преобладали дуб, клен, ясень, липа, пихта и редко кедр» [1]. Соотношение хвойных и лиственных менялось в зависимости от местоположения – лиственные преобладали на низменных местах и приморских склонах, а хвойные – на более высоких элементах рельефа. Главный хребет полуострова был полностью покрыт лесом с преобладанием хвойных, наиболее широко была распространена пихта цельнолистная [1]. Об этом свидетельствуют и исследования лесов, проведенные в 1886 г. Для лесов западного побережья полуострова и в центральной части полуострова отмечалось доминирование пихты над кедром [9]. В пределах России чернопихтово-широколиственные леса развиты только на крайнем юге Приморского края, где проходит северо-восточная граница их ареала в Восточной Азии. Основной ареал находится за пределами России – на сопредельных территориях северо-восточной части Китая и Корейского полуострова. Леса этой формации характеризуются высокими показателями видового богатства и совершенно справедливо включены в список редких растительных сообществ Приморского края [8]. Несмотря на длительное и разнообразное влияние человека на леса полуострова, чернопихтово-широколиственные леса здесь характеризуются сложной вертикальной структурой, большой степенью горизонтальной неоднородности и высокой видовой насыщенностью.
Растительность на полуострове за последние полтора столетия претерпела наиболее значительные изменения, а на застроенных участках полностью уничтожена. Широколиственные и хвойно-широколиственные леса сохранились лишь в северной и восточной частях полуострова. Здесь выделяются условно-коренные и производные сообщества, формирующиеся на месте коренных лесов [15]. В настоящее время на полуострове широко распространены вторичные дубовые леса, широколиственные леса с грабом, липами, кленами и дубом монгольским; на хребте Богатая Грива сохранились участки грабово-широколиственно-кедровых лесов с цельнолистной пихтой и чернопихтово-широколиственные леса [5]. Луга и болота занимают менее 5 % площади. По геоботаническому районированию Дальнего Востока [6] рассматриваемая территория относится к Восточно-Азиатской хвойно-широколиственной области.
История развития ландшафтов этой территории представляет большой интерес с точки зрения развития коренных растительных сообществ при разнонаправленных короткопериодных изменениях климата, и выяснения соотношения природных и антропогенных факторов в их преобразовании.
Полуостров Муравьева-Амурского входит в климатическую область муссонов умеренных широт, которая характеризуется контрастной сменой сезонной циркуляции воздушных масс и гидротермических параметров. По количеству осадков территория относится к зоне достаточного увлажнения. Среднегодовое количество осадков – 700–800 мм, из них до 80 % выпадает в теплый период года (апрель – сентябрь). Среднегодовые температуры составляют 3–6 °С, сумма активных температур 2800–2900 °С, продолжительность вегетационного периода до 190 дней.
Для палеоландшафтных реконструкций была выбрана бухта Муравьиная, расположенная в кутовой части Уссурийского залива (рис. 1). В бухту впадают реки Шкотовка, Артёмовка, в устьях которых расположены эстуарии, и более мелкие речки и ручьи. Вода в бухте опреснена, глубины невелики (2–5 м). Берега бухты преимущественно аккумулятивные, низменные и заболоченные, на западном берегу хорошо выражена морская терраса (высота 2–2,5 м). На западном берегу расположено озеро Черепашье (420х500 м), в него впадает безымянный ручей, нижняя часть долины которого, как и берега озера, сильно заболочена. Окружающий рельеф преимущественно низкогорный (200–400 м над уровнем моря), сильно расчлененный с относительными превышениями до 100–150 м; крутизна склонов колеблется от 2 ° до 30 °. В 3 км к юго-западу от озера Черепашье находится г. Пионерская (абс. высота 255 м), в 10 км к западу – самая высокая на полуострове вершина с абс. высотой 474 м.
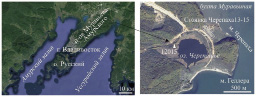
Рис. 1. Схема района работ
Современная растительность прилегающих к озеру склонов представлена вторичными, преимущественно дубовыми лесами из дуба монгольского (Quercus mongolica Fisch. ex Ledeb.) при участии лип амурской и маньчжурской (Tilia amurensis Rupr., T. mandshurica Rupr.), березы даурской (Betula davurica Pall.), ореха маньчжурского (Juglans mandshurica Maxim.), кленов маньчжурского и мелколистного (Acer mandshuricum Maxim., A. mono Maxim.) при обилии в подчиненных ярусах граба сердцелистного (Carpinus cordata Blume), клена ложнозибольдова (A. pseudosieboldianum (Pax) Kom.). Единичные хвойные сохранились преимущественно в трудно доступных верхних частях склонов, преимущественно это пихта цельнолистная (Abies holophylla Maxim.), реже – кедр корейский (Pinus koraiensis Siebold et Zucc.). Пихта белокорая (Abies nephrolepis (Trautv.) Maxim.) на полуострове большая редкость. В нижнем течении ручья, впадающего в озеро Черепашье, представлены сырые ольховые леса из ольхи волосистой (Alnus hirsuta (Spach) Fisch. ex Rupr.). Морская терраса занята приморскими лугами на дерново-луговых почвах.
На аккумулятивной форме, отделяющей северный берег озера от акватории бухты, расположен археологический памятник раннего железного века – янковская культура (2,8–2,0 тыс. л.н.) (устное сообщение с.н.с. ООО «НПЦ ИКЭ» С.С. Малкова). Мыс Черепаха защищает от ветра этот участок берега, что создает благоприятный микроклимат. В ходе археологических раскопок 2015 г. здесь были обнаружены котлованы жилищ, хозяйственные ямы, раковинные кучи и несколько человеческих захоронений.
Целью работы является восстановление природных условий п-ова Муравьев-Амурский в течение среднего-позднего голоцена, в том числе и во время возникновения и существования на побережье бухты Муравьиная поселения янковской культуры.
Материалы и методы исследования
На низинном тростниковом болоте, где доминирует тростник южный (Phragmites australis (Cav.) Trin. ex Steud.) при участии тростника высочайшего (P. altissimus (Benth.) Nabille), в 70 м от западного берега озера Черепашье (630 м от уреза моря, абс. высота около 1 м) с помощью ручного бура была взята колонка разнофациальных отложений (мощность 2,2 м). Образцы на спорово-пыльцевой и диатомовый анализы были отобраны с шагом 5 см и обработаны по стандартным методикам. Возрастная привязка палеоландшафтных смен сделана на основе радиоуглеродного датирования, выполненного в СПбГУ (таблица).
|
Лаб. номер |
Номер образца |
Интервал, м |
Материал |
Радиоуглеродный возраст, лет |
Календарный возраст cal BP, лет |
|
ЛУ-8024 |
1/12015 |
0,45–0,50 |
торф |
820 ± 80 |
760 ± 80 |
|
ЛУ-8023 |
2/12015 |
0,70–0,75 |
торф |
1080 ± 90 |
1010 ± 110 |
|
ЛУ-8025 |
3/12015 |
0,95–1,00 |
торф |
2010 ± 90 |
1980 ± 120 |
|
ЛУ-8026 |
4/12015 |
1,15–1,20 |
торф |
1610 ± 80 |
1500 ± 90 |
|
D-AMS 16826 |
44/12015 |
2,15–2,20 |
раковины |
5550 ± 21 |
6346 ± 32 |
Примечание: значения календарного возраста приведены на основании калибровочной программы «OxCal 4.2», калибровочная кривая «IntCal 13» (https://c14.arch.ox.ac.uk).
Образцы торфа были предварительно очищены от карбонатов и посторонних гуминовых кислот путем последовательной обработки образцов 2 % растворами HCl и NaOH. Возраст отложений из нижней части разреза определен с помощью AMS-датирования в лаборатории «DirectAMS», г. Ботелл (Bothell), США. Калибровка радиоуглеродных дат в календарные сделана с помощью программы OxCal 4.2 (https://c14.arch.ox.ac.uk). При описании диатомовых комплексов и палинозон приведены календарные возраста, полученные с применением возрастной модели по программе Bacon 2 и с программной оболочкой R [17]. Рассчитанный радиоуглеродный возраст приведен в соответствие с полученными по модели калиброванными датировками [20]. Для терригенных отложений возраст определялся с допущением одинаковых скоростей осадконакопления на основе датирования раковин моллюсков, залегающих в основании лагунных отложений. Поскольку скорости осадконакопления могли быть непостоянными, хронологические данные, полученные для этой части разреза, рассматриваются как приблизительные.
Во время раскопок в 2015 г. археологического памятника Черепаха-13 на спорово-пыльцевой анализ были отобраны пробы из заполнения хозяйственных построек, развалов сосудов и мест погребений.
Результаты исследования и их обсуждение
В основании изученного разреза (2,05–2,20 м) залегает голубовато-серый ил с обломками и единичными целыми раковинами устриц (Crassostrea gigas); выше он переходит в оливковый алевро-пелитовый ил (1,20–2,05 м), в нижней части включающий примесь разнозернистого песка. Накопление илов началось в конце атлантического периода голоцена (таблица) – с учетом эффекта резервуара, равном 400 лет [18], около 5150 14С л.н. (5950 кал л.н.). Верхнюю часть разреза слагает темно-коричневый умеренно и плохо разложившийся травяной торф (0–1,20 м), образование которого началось в конце суббореального периода. Датировка ЛУ-8026 из нижней части торфяника рассматривается как омоложенная. По скоростям торфонакопления возраст торфяника оценивается около 2750 14С лет (2490 кал. л.н.). В торфянике на глубине 0,83 м обнаружено гнездо вулканического пепла B-Tm извержения влк. Байтоушань в 969 г. 14С-дата ЛУ-8023 из вышележащего торфа показывает, что это гнездо было «затянуто» в нижележащие слои по вертикально ориентированным корням болотных растений. На начальном этапе скорости торфонакопления были низкими (0,26 мм/год), в малый оптимум голоцена они возросли (до 1 мм/год) и несколько снизились в малый ледниковый период (0,66 мм/год) (рис. 2).
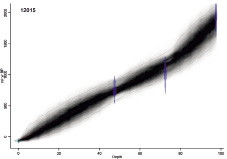
Рис. 2. Возрастная модель накопления отложений на берегу оз. Черепашье в среднем-позднем голоцене
Список радиоуглеродных дат из разреза голоценовых отложений на побережье бух. Муравьиная
Диатомовый анализ. В отложениях разреза обнаружено 116 видов и разновидностей пресноводных и 11 видов солоноватоводных и морских диатомовых водорослей. Изменение содержания видов диатомей по разрезу позволило выделить 5 комплексов, отражающих смену обстановок осадконакопления (рис. 3).

Рис. 3. Распределение диатомей в отложениях разреза 12015 на берегу оз. Черепашье, бух. Муравьиная. Экологическая характеристика диатомей – место обитания: 1 – планктон, 2 – донные, 3 – обрастания; соленость: 1 – галофобы, 2 – индифференты, 3 – галофилы, 4 – мезогалобы; рН: 1 – ацидофилы, 2 – циркумнейтральные, 3 – алкалифилы; биогеография: 1 – арктические, 2 – бореальные, 3 – космополиты. 1 – торф, 2 – алевро-пелитовые отложения, 3 – песок, 4 – раковины и их обломки
Комплекс 1 (инт. 1,90–2,20 м, 5150–4360 14С л.н., 5950–4910 кал. л.н.). В составе диатомей преобладают пресноводные виды обрастания: Pseudostaurosira elliptica, Fragilaria construens, Staurosira construens var. binodis, Pseudostaurosira subsalina, Staurosirella pinnata (в сумме до 89,4 %), характерные для стоячих вод. Здесь же отмечено высокое содержание бентосных морских и солоноватоводных, наиболее часто встречаются Pinnunavis jarrensis (до 22,8 %), Diploneis interrupta, Amphora commutata, присутствует морской вид Terpsinoe americana, характерный для хорошо прогреваемых вод. По отношению к рН доминируют алкалифилы, по отношению к солености – индифференты и галофилы. В целом комплекс свидетельствует о существовании на побережье бух. Муравьиная в конце атлантического периода сильно распресненной мелководной лагуны, имеющей хорошую связь с морем.
Комплекс 2 (инт. 1,55–1,90 м, 4360–3480 14С л.н., 4910–3700 кал. л.н.) характеризуется резким увеличением количества морских и солоноватоводных видов диатомей (до 74,6 %). Доминирует солоноватоводный Pinnunavis jarrensis, субдоминантами являются Campylodiscus echeneis и Terpsinoe americana (до 3,3 %). Заметно сокращается количество видов обрастания (до 55 %), что может свидетельствовать о разрушении сообществ водной растительности из–за увеличения площади и глубины лагуны. Все вместе это говорит о некотором повышении уровня моря в условиях потепления.
Комплекс 3 (инт. 1,10–1,55 м, 3480–2230 14С л.н., 3700–2200 кал. л.н.) в целом близок комплексу 1. Содержание морских и солоноватоводных видов снижается до 12–16 %, лагуна становится более закрытой и более распресненной. Здесь преобладают виды обрастания (до 90,5 %): Pseudostaurosira elliptica, Fragilaria construens, Staurosira construens var. binodis, Pseudostaurosira subsalina, Staurosirella pinnata, характерные для стоячих вод. По отношению к солености преобладают индифференты, по отношению к рН – алкалифилы. Количество морских и солоноватоводных видов сокращается до 19,5 %, возможно, уменьшается связь с морем. Комплекс свидетельствует о существовании мелководной лагуны с заболоченными берегами.
Комплекс 4 (инт. 0,95–1,10 м, 2230–1820 14С л.н., 2200–1760 кал. л.н.). Сокращается число видов-обрастателей, совсем исчезают виды рода Staurosira, увеличивается число донных видов (до 61 %). Доминируют солоноватоводный бентосный Diploneis interrupta, и характерные для пресных и слегка солоноватых водоемов D. oblongella, D. ovalis, из пресноводных – Hantzschia amphioxis, Caloneis bacillum, также резко увеличивается количество видов рода Eunotia, что свидетельствует о сокращении площади озера-лагуны и развитии болотных обстановок по его обрамлению, что, скорее всего, связано с регрессией в начале субатлантика (14С-дата 2010 ± 90 л., 1980 ± 120 кал. л., ЛУ-8025). Значительно возрастает количество бореальных диатомей – условия стали менее тепловодными.
Комплекс 5 (инт. 0–0,95 м, последние 1820 14С лет, 1760 кал. л.н.). Присутствуют только пресноводные диатомеи, доминируют донные виды (до 82 %): Diploneis ovalis, D. oblongella, Hantzschia amphioxis, Caloneis bacillum, появляются виды родов Navicula, Nitzschia, Pinnularia. Из обрастателей появляются Eunotia glacialis, E. praerupta, характерные для болотных обстановок, и Fragilaria nitzschioides. Увеличение доли таких видов, как Navicula radiosa, Nitzschia sinuata var. delognei в инт. 0–0,15 м, могут показывать усиление влияния вод ручья последние 200 лет. Исчезновение морских и солоноватоводных видов свидетельствует о прекращении связи водоема с морем. В целом в этой части разреза увеличивается число арктобореальных видов (до 10,5 %), в то же время в интервале 0,50–0,60 м количество арктобореальных видов резко уменьшается (1,4–4,7 %). В целом комплекс отвечает развитию болотных обстановок по обрамлению мелководного пресного водоема в последние 2 тыс. лет. В торфянике в отдельных пробах найдены морские сублиторальные бентосные диатомеи Tryblionella granulata, Campylodiscus echeneis и солоноватоводный Diploneis interrupta.
Спорово-пыльцевой анализ. В нижней части разреза в лагунных илах в составе палиноспектров преобладает пыльца древесных, что отвечает развитию сомкнутых лесов на окружающих склонах, количество пыльцы травянистых не превышает 34,6 % (рис. 4). В нижней части органогенных отложений увеличивается количество пыльцы трав и кустарничков (до 43,9 %), в верхней части разреза резко увеличивается количество спор (до 89,1 %), а в кровле разреза – пыльцы трав (до 61,2 %), что отвечает развитию кустарниковых группировок и лугов на барьерных формах и существованию заболоченных участков около лагуны. Распределение пыльцы и спор по разрезу позволяет выделить 7 палинозон.

Рис. 4. Спорово-пыльцевая диаграмма отложений разреза 12015 на берегу оз. Черепашье, бух. Муравьиная (начало)

Рис. 4. Спорово-пыльцевая диаграмма отложений разреза 12015 на берегу оз. Черепашье, бух. Муравьиная (окончание)
Палинозона 1 (2,0–2,2 м, 5150–4480 14С л.н., 5950–5250 кал. л.н.) отвечает распространению на побережье и окружающих склонах кедрово-дубовых лесов с участием граба, липы, берез. В подлеске были распространены лещина (Corylus), виноград (Vitis), сирень (Syringa), на осветленных участках – аралия (Aralia). Ильм (Ulmus), орех, ясень (Fraxinus) и кедр корейский могли расти по долинам водотоков и в нижних частях склонов. Пыльца темнохвойных пород и Pinus s/g Diploxylon встречена в небольших количествах. В это время пихта цельнолистная уже принимала участие в лесной растительности полуострова. Пыльца ели (Picea), скорее всего, занесена с удаленных горных хребтов. Пыльца сосны густоцветковой (Pinus densiflora Siebold et Zucc.) также могла быть занесена ветром. Аккумулятивные формы были покрыты полынно-разнотравными сообществами. В обрамлении лагуны были распространены осоковые ассоциации, заросли рогоза (Typha).
Палинозона 2 (1,25–2,0 м, 4480–2460 14С л.н., 5250–2660 кал. л.н.) характеризуется большим содержанием пыльцы широколиственных и увеличением их разнообразия. В палиноспектрах зафиксировано максимальное содержание пыльцы широколиственных, в том числе граба (до 9 %). На склонах в обрамлении бухты существовали полидоминантные широколиственные леса с участием дуба, граба, ильма, ореха, диморфанта (Kalopanax), ясеня, бархата (Phellodendron), клена, липы, березы, кедра корейского. В подлеске участвовали лещина, на осветленных участках – аралия и калина (Viburnum). Выше по склонам были распространены кедрово-широколиственные леса. Количество пыльцы кедра вверх по разрезу увеличивается, что свидетельствует о постепенном увеличении его роли в лесной растительности полуострова. Присутствие пыльцы можжевельника (Juniperus) свидетельствует, что отдельные куртины можжевельника, возможно, были на скальных участках мысов. Среди трав на аккумулятивных формах были широко представлены полынь (Artemisia) и разнотравье, на низменных берегах лагуны существовали заболоченные участки с осоковыми и вейниковыми группировками. В верхней части интервала резко увеличивается содержание пыльцы рогоза.
Палинозона 3 (1,15–1,25 м, 2460–2320 14С л.н., 2660–2340 кал. л.н.) отличается увеличением содержания пыльцы хвойных (Pinus s/g Haploxylon – до 37 %, Picea – до 14 %, Abies – 2,7 %) и сокращением количества и разнообразия пыльцы широколиственных (до 27 %). По обрамлению бух. Муравьиной были распространены широколиственно-кедровые леса, на вершинах появились небольшие массивы кедрово-елово-широколиственных лесов, насыщенных неморальными элементами. В настоящее время такие леса известны в Уссурийском заповеднике, где занимают высотный интервал от 400–500 до 600–650 м. В составе этих лесов присутствуют ель аянская (Picea ajanensis (Lind. et Gord.) Fisch. ex Carr.), кедр корейский, пихта белокорая, единично пихта цельнолистная, липы (амурская и Таке) (Tilia amurensis, T. taquetii C.K. Schneid.), клен мелколистный, диморфант), береза желтая (B. costata Trautv.) [16]. Среди трав резко сократилось содержание пыльцы полыни.
Палинозона 4 (1,05–1,15 м, 2320–2090 14С л.н., 2340–2050 кал. л.н.) характеризуется увеличением содержания пыльцы широколиственных: количество пыльцы дуба достигает 44 %, пыльцы других широколиственных – 15 %. Содержание пыльцы кедра корейского падает до 15 %, а ели – до 3 %. Палиноспектры отражают широкое развитие на побережье бухты и в ее обрамлении кедрово–дубовых лесов со значительной примесью граба (Carpinus – до 10 %) и берез (в сумме до 21 %). Увеличение содержания пыльцы трав, представленных главным образом осоками (Carex), отражает активное заболачивание берегов лагуны и нижней части долин водотоков.
Палинозона 5 (0,70–1,05 м, 2090–1050 14С л.н., 2050–1100 кал. л.н.) характеризуется сокращением содержания пыльцы широколиственных пород (до 27 %) и увеличением хвойных (Pinus s/g Haploxylon до 31 %, Pinus s/g Diploxylon до 31 %, Picea до 15 %). В обрамлении бухты росли широколиственно-кедровые леса, на высоких уровнях рельефа расширились площади кедрово-елово-широколиственных лесов, возможно, отдельные скалистые участки на полуострове и берегу моря занимали небольшие рощи из сосны густоцветковой. Увеличение содержания пыльцы ольхи говорит о том, что на побережье и в долине ручья стали распространяться ольховые леса с папоротниковым покровом. Среди пыльцы трав доминируют осоки. Вверх по разрезу сокращается количество пыльцы рогоза. Появляется пыльца рдеста (Potamogeton), характерного для мелководий стоячих и слабо проточных водоемов. В палиноспектрах увеличивается доля спор, представленных в основном папоротниками. В напочвенном покрове в ближайшем окружении лагуны были широко распространены зеленые мхи.
Палинозона 6 (0,25–0,70 м, 1050–320 14С л.н., 1100–370 кал. л.н.) характеризуется резким увеличением содержания пыльцы ольхи (до 57 %). Вокруг озера расширялись ассоциации с ольхой и осоками. На склонах были распространены кедрово-широколиственные леса, разнообразие широколиственных таксонов сокращается. Вероятно, сократились участки, занятые соснами Pinus s/g Diploxylon.
Палинозона 7 (0,00–0,25 м, последние 320 14С л.н., 370 кал. л.н.). В общем составе спектров наблюдается уменьшение доли древесной пыльцы (< 4 %) и пыльцы трав, увеличивается количество спор, представленных в основном папоротниками. Это может свидетельствовать об увеличении открытых участков на склонах, возможно, за счет пожаров. Среди древесных возрастает роль кедра корейского (44 % в группе древесной пыльцы), роль широколиственных сокращалась. В составе болотной растительности по берегам озера увеличивается участие осок и сфагновых мхов. Присутствие пыльцы хвойника (Ephedra) можно объяснить ее заносом с песчаных аккумулятивных форм, распространенных в кутовой части Уссурийского залива. В кровле торфяника содержание пыльцы дуба составляет 46 %, появляется пыльца граба, увеличивается доля пыльцы березы, резко сокращается содержание пыльцы хвойных. Такие изменения могли быть связаны с антропогенной трансформацией растительности и в какой-то мере с потеплением после малого ледникового периода.
Спорово-пыльцевые данные для археологического памятника Черепаха-13. Спорово-пыльцевые спектры характеризуются преобладанием пыльцы трав (до 91 %), что отвечает локальной растительности на аккумулятивных формах, где было расположено поселение (рис. 5).

Рис. 5. Спорово-пыльцевая диаграмма культурного слоя археологического памятника Черепаха-13
Доминирует пыльца полыни (до 70 %) и других астровых (Asteraceae) (до 14 %), маревых (Chenopodiaceae) (до 24 %), лютиковых (Ranunculaceae) (до 24 %), злаковых (Poaceae) (до 9 %), присутствует пыльца осоковых (Cyperaceae), зонтичных (Apiaceae), розовых (Rosaceae) (репяшок мелкобороздчатый Agrimonia striata Michx. и кровохлебка Sanguisorba), валериановых (Valerianaceae), бобовых (Fabaceae), спорышовых (Polygonaceae), норичниковых (Scrophulariaceae), гвоздиковых (Сaryophyllaceae), яснотковых (Lamiaceae), капустовых (Brassicaceae), крапивы (Urtica) и подмаренника (Galium). Особенно много пыльцы полыни (92 %) обнаружено в образце с пола жилища. В одной из хозяйственных ям была найдена пыльца культурных злаков (3 %), скорее всего проса. В двух образцах среди пыльцы древесных отмечена пыльца ели. Из непыльцевых палиноморф найдены споры гриба Gelasinospora – индикатора пожаров (мог поселиться на месте старого очага), и споры гриба Glomus – индикатора почвенной эрозии. Встречены зеленые водоросли Zygnema-type, характерные для стоячих вод, которые могли быть занесены сюда древним человеком. В жилищах встречены створки морских диатомовых водорослей (Arachnoidiscus ehrenbergii), которые могли быть принесены с грунтом или продуктами моря. Также были обнаружены яйца тихоходок рода Macrobiotus, которые обычно многочисленны на мхах, яйца коловраток (Rotaria), обитающих как в морских, так и в пресноводных условиях и на влажных почвах. В могильниках присутствуют споры грибов Glomus и Meliola, зеленые пресноводные водоросли Spyrogyra, фрагменты вислокрылок или ручейников. Из древесных таксонов в погребениях встречена только пыльца березы.
Комплексное изучение разреза разнофациальных отложений на побережье бух. Муравьиной позволяет восстановить ход развития ландшафтов п-ова Муравьева-Амурского за последние 5150 14С лет, 5946 кал. лет (рис. 6).

Рис. 6. Реконструкция смены обстановок осадконакопления на побережье бух. Муравьиная и изменения ландшафтов п-ова Муравьева-Амурского в среднем-позднем голоцене
Неморальные леса существуют здесь как минимум с оптимума голоцена. В завершающую фазу атлантика были распространены полидоминантные широколиственные леса с преобладанием дуба. Участие сосны корейской в составе лесной растительности было незначительным. Аналогичные леса занимали весь полуостров и о. Русский [10] и тянулись полосой вдоль побережья Уссурийского залива [3]. Пихта цельнолистная принимала участие в лесной растительности полуострова в конце атлантика, а возможно, появилась ранее, как и на западном побережье Амурского залива [11].
На пике голоценовой трансгрессии (около 6000 14С л.н.), когда уровень моря был выше современного на 2–3 м [7], на месте озера Черепашье существовала мелководная распресненная лагуна с устричным рифом. Оптимальные условия для обитания Crassostrea gigas – глубина до 3–5 м, температура в летний период – не менее 18 °С, соленость воды – около 25–28 ‰ [14], но устрицы могут жить в распресненных условиях и кратковременно выдерживают понижение солености до 5 ‰ [19]. Устричники в заливе Петра Великого были широко распространены в оптимальную фазу голоцена [14]. В изученном разрезе, скорее всего, вскрыты морские отложения, которые накапливались в завершающую стадию трансгрессии. Климатические условия были близки к современным. В регрессию на границе атлантик – суббореал осадконакопление в лагуне не прерывалось, в осадке появляется примесь разнозернистого песка.
В суббореальный максимум в составе лесной растительности полуострова увеличивалось участие термофилов, в первую очередь граба. Это говорит о теплых климатических условиях со среднегодовой температурой свыше + 6 °С и с количеством осадков до 1000–1200 мм [7]. Ограниченные площади занимали чернопихтово-широколиственные леса. Это потепление ярко проявилось и на о. Русский [10], где были распространены широколиственные леса с преобладанием дуба и граба. Полидоминантные широколиственные леса были распространены вплоть до Шкотовского плато [13]. Такие ландшафты на побережье бух. Муравьиная были достаточно стабильными длительное время – 4480–2460 14С л.н. (5250–2660 кал. л.н.), малоамплитудная климатическая ритмика, оказавшая влияние на развитие ландшафтов более открытого побережья (о. Русский) и в горных районах Приморья, здесь не проявилась. Постепенно в течение 860 лет (3290–2460 14С л.н., 3520–2660 кал. л.н.) в лесной растительности увеличивалось участие сосны корейской. Резкое увеличение в палиноспектрах содержания пыльцы полыни может свидетельствовать о ее распространении на аккумулятивной форме и прибрежных скалах около 3000–2700 14С л.н., 3180–2830 кал. л.н.
На пике суббореальной трансгрессии лагуна имела хорошую связь с морем, увеличивалась глубина и соленость водоема. При снижении уровня моря во второй половине суббореала лагуна стала более закрытой и распресненной. В конце этой стадии на западном берегу лагуны происходит смена терригенного осадконакопления на биогенное. По-видимому, около 2440 14С л.н., 2490 кал. л.н. началось активное заболачивание и торфонакопление.
Предположительно именно в это время на побережье бух. Муравьиная появляются поселения раннего железного века, связанные с использованием морских ресурсов. Палиноспектры из образцов с археологической стоянки характеризуются доминированием пыльцы трав, что отвечает развитию приморских лугов вблизи поселения. Значительное количество спор гриба Glomus говорит о распространении почвенной эрозии, также в результате хозяйственной деятельности человека. Об активном использовании древними людьми морских ресурсов свидетельствуют находки зеленых Zygnema-type и диатомовых водорослей. Находки пыльцы культурных злаков в хозяйственной яме указывают на их употребление в пищу, а не на культивирование.
Похолодание на границе суббореал – субатлантик, имевшее глобальный характер [7], проявилось на побережье бух. Муравьиная около 2460–2320 14С л.н., 2660–2340 кал. л.н. и вызвало существенные изменения в растительности окружающих склонов. На полуострове распространились широколиственно-кедровые леса, в привершинных участках хребтов появились кедрово-елово-широколиственные леса с неморальными элементами. В это время произошла экспансия темнохвойных лесов на Шкотовском плато, краевые части которого также занимали широколиственно-кедровые леса [13].
В палиноспектрах из основания торфяника хорошо фиксируется раннесубатлантическое потепление (около 2320–2090 14С л.н., 2340–2050 кал. л.н.), когда на побережье бухты получили распространение кедрово-дубовые леса с участием граба и березы (Betula sect. Costatae). Это потепление проявилось и на других участках побережья Южного Приморья; климатические условия были близки к современным или были несколько прохладнее [11].
Широколиственно-кедровые леса в горном обрамлении бух. Муравьиная были достаточно стабильными длительный промежуток времени (2090–1050 14С л.н., 2050–1100 кал. л.н.). Роль кедра корейского увеличилась в более прохладных условиях. Присутствие большого количества пыльцы Pinus s/g Diploxylon свидетельствует, что сосна густоцветковая, скорее всего, участвовала в растительности побережья и крутых каменистых склонах. В настоящее время небольшие участки, занятые сосняками, сохранились в долине р. Артемовка. На полуострове единичные экземпляры сосны густоцветковой сохранились на водоразделах рек Пионерская (Седанка) и Богатая [15].
Снижение уровня моря в первой половине субатлантика [7] привело к закрытию лагуны и превращению ее в пресноводное озеро около 1820 14С л.н., 1760 кал. л.н. Именно с этого времени, когда, по-видимому, произошло уменьшению площади озера и осушение поверхности его берегов, вокруг озера и в нижней части долины ручья расширились площади ольховников с папоротниковым покровом. Более дробная климатическая ритмика, выделенная на открытом побережье Южного Приморья [11], на изученном участке берега не была выявлена.
Еще более широкое распространение вокруг озера ольховые леса получили в малый оптимум голоцена и первую половину малого ледникового периода. Начиная с малого оптимума в обрамлении бухты стали доминировать кедрово-широколиственные леса более бедного состава по сравнению с ранее существовавшей лесной растительностью. Потепление не было ярко проявлено, но «теплый сигнал» выделяется по увеличению количества пыльцы дуба, липы и снижению содержания арктобореальных видов диатомей около 950–790 кал. л.н. Возможно, в изученном разрезе фиксируется потепление конца XII века, которое ярко проявилось в Китае [4].
Снижение температуры и ухудшение климатических условий во вторую половину малого ледникового периода (XVI–XIX вв.) привело к существенным ландшафтным изменениям. На побережье увеличилась площадь безлесных участков, а в горах в лесной растительности увеличилась роль кедра корейского. В составе широколиственно-кедровых лесов уменьшалось участие неморальных элементов. Возможно, расширились участки неморальных ельников на верхних уровнях рельефа. Вероятно, на побережье могли существовать рощи сосны густоцветковой. Вокруг озера преобладали сфагново-осоковые ассоциации. О холодных условиях свидетельствует увеличение содержания арктобореальных видов диатомей. Преобладание хвойно-широколиственных лесов с кедром корейским и сосной густоцветковой в малый ледниковый период отмечается также и по данным спорово-пыльцевого анализа почвенных профилей на восточном побережье п-ова Муравьева-Амурского [12]. Похолодание малого ледникового периода также зафиксировано и в отложениях на о. Русский [10]. Роль дуба в ХХ веке значительно увеличилась в связи с антропогенным влиянием. Палиноспектры свидетельствуют о значительной трансформации коренной растительности, обеднении ее состава, резком уменьшении роли хвойных.
Находки морских диатомей в торфе, который формировался на берегу пресного озера, по-видимому, могут являться свидетельством прохождения цунами. Возраст этих событий оценивается около 1170, 1000, 500 кал. л.н. Следы близких по возрасту сильных цунами обнаружены на побережье Восточного Приморья [2].
Выводы
Развитие ландшафтов в обрамлении бух. Муравьиной в среднем-позднем голоцене достаточно четко коррелируется с изменениями климата и колебаниями уровня моря. Трансгрессии и регрессии среднего-позднего голоцена существенно меняли ситуацию на побережье и были основными факторами, определяющими формирование аккумулятивных форм и эволюцию лагуны, прошедшую пять стадий развития от водоема в разной степени связанного с морем до пресноводного озера.
Полидоминантные широколиственные леса, которые были распространены на п-ове Муравьева-Амурского в оптимум голоцена, в начале позднего голоцена постепенно заместились кедрово-широколиственными. Роль хвойных сильно возросла во время похолодания на границе суббореал – субатлантик. Биотические компоненты ландшафтов быстро отреагировали на раннесубатлантическое потепление, когда в лесной растительности возросла роль неморальных элементов. Широколиственно-кедровые леса в горном обрамлении бух. Муравьиная были стабильными длительный промежуток времени и еще существовали в начале малого оптимума голоцена. Это потепление не было ярко проявлено, но со второй половины малого оптимума в обрамлении бухты стали распространены кедрово-широколиственные леса более бедного состава по сравнению с ранее существовавшей лесной растительностью. Широкое развитие ольховников на побережье связано со снижением уровня моря в первую половину субатлантика. Существенные ландшафтные изменения произошли во второй половине малого ледникового периода: в лесной растительности увеличилась роль хвойных, на побережье увеличилась площадь безлесных участков. Значительная антропогенная трансформация лесной растительности с образованием вторичных дубняков произошла в ХХ веке.
Полученные данные позволили также восстановить природную среду в период возникновения и существования на побережье бухты Муравьиная поселений янковской культуры (2,8–2,0 тыс. кал. л.н.). Стоянка возникла, когда климатические условия были теплее современных, уровень моря был выше современного, на побережье были развиты полидоминантные широколиственные леса с участием кедра корейского. Во время существования поселения произошли климатические изменения в сторону похолодания, сопровождавшиеся падением уровня моря, это ухудшило условия для проживания здесь людей и, возможно, привело к полному прекращению существования поселения.
Работа выполнена по грантам РФФИ 15-05-00171, 15-05-00179.
Библиографическая ссылка
Лящевская М.С., Макарова Т.Р., Разжигаева Н.Г., Ганзей Л.А., Кудрявцева Е.П., Паничев А.М., Арсланов Х.А., Максимов Ф.Е., Петров А.Ю. РАЗВИТИЕ ЛАНДШАФТОВ ПОЛУОСТРОВА МУРАВЬЕВА-АМУРСКОГО В СРЕДНЕМ-ПОЗДНЕМ ГОЛОЦЕНЕ ПО ДАННЫМ ИЗУЧЕНИЯ ОТЛОЖЕНИЙ ПОБЕРЕЖЬЯ БУХТЫ МУРАВЬИНАЯ (ЮЖНОЕ ПРИМОРЬЕ) // Успехи современного естествознания. 2017. № 2. С. 110-122;URL: https://natural-sciences.ru/en/article/view?id=36372 (дата обращения: 17.07.2026).