


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
IN VITRO DIGESTIBILITY OF STORAGE ENDOSPERM PROTEINS OF TRANSGENIC SORGHUM PLANTS CARRYING GENETIC CONSTRUCT FOR SILENCING OF THE GAMMA-KAFIRIN GENE

Создание сортов и гибридов, обладающих нужными для человека признаками и свойствами, является основной целью селекции растений. С развитием методов генетической инженерии возможности селекционеров и генетиков по созданию новых сортов и гибридов значительно расширились. Одним из перспективных направлений использования генетической инженерии в селекции является улучшение питательной ценности возделываемых культур, а именно изменение содержания белка, крахмала, незаменимых аминокислот, подавление синтеза аллергенов и различных веществ, обладающих антипитательными свойствами [1, 2].
Особенно актуальны эти работы для сорго – высокоурожайной засухоустойчивой злаковой культуры. Сорго входит в число пяти наиболее широко возделываемых злаковых культур мирового сельскохозяйственного производства. С потеплением климата и увеличением посевных площадей, подвергающихся воздействию засухи, эта культура будет получать все большее распространение. В то же время, по сравнению с другими видами злаков, большинство ныне возделываемых сортов и гибридов сорго имеют более низкую питательную ценность [3].
Одной из причин сравнительно низкой питательной ценности зерна сорго является устойчивость его запасных белков (кафиринов) к расщеплению протеазами. При этом считается, что наибольшей устойчивостью к протеолитическому расщеплению обладает гамма-кафирин, занимающий периферийное положение в белковых тельцах клеток эндосперма и тем самым блокирующий доступ протеаз к альфа-кафиринам, располагающимся внутри белковых телец [4, 5].
Развитие методов генетической инженерии открывает возможность решения этой проблемы с помощью технологии РНК-интерференции, позволяющей целенаправленно «выключать» экспрессию отдельных генов. В последние годы технология РНК-интерференции стала широко использоваться для изменения состава запасных белков и крахмала у разных видов злаков, в том числе и у сорго [6–8].
Нами ранее сообщалось о получении трансгенных растений сорго, несущих генетическую конструкцию, способную к индукции РНК-сайленсинга гена гамма-кафирина [9]. В данной статье представлены результаты исследования перевариваемости запасных белков в системе in vitro у потомства полученных трансгенных растений.
Материалы и методы исследования
Исследовали потомство (поколения Т3–T4) трансгенных растений, полученных ранее в опыте по генетической трансформации незрелых зародышей линии Желтозерное-10 (Ж10) с использованием штамма Agrobacterium tumefaciens GV3101, несущего бинарный вектор pNRKafSil, созданный в Центре биоинженерии РАН [9]. Данный вектор содержит в составе Т-ДНК селективный маркерный ген bar под контролем nos-промотора и целевую вставку, представляющую собой инвертированный повтор последовательности внутренней области гена гамма-кафирина сорго, разделенный последовательностью интрона гена ubi1 кукурузы, под контролем 35S-промотора.
У зерновок, завязавшихся на растениях, исследовали текстуру эндосперма (рассекая скальпелем зерновку на две половины), а также перевариваемость запасных белков. Для изучения перевариваемости белков использовали метод обработки муки пепсином в условиях in vitro, широко применявшийся ранее в ряде исследований [10–12]. Мука (20 мг), полученная от цельносмолотых зерновок трансгенных растений и исходной нетрансгенной линии Ж10, подвергалась обработке 0,15 % раствором пепсина (P7000 Sigma-Aldrich) в 0,1 M калий-фосфатном буфере (pH 2,0) в течение 2 час при 37 °C [10]. Контрольные образцы инкубировали в калий-фосфатном буфере без добавления пепсина при тех же условиях.
Для количественной оценки перевариваемости белков использовали информативный метод, основанный на SDS-электрофорезе в полиакриламидном геле (ПААГ) c последующей денситометрией полученных спектров [12]. Для этой цели образцы подвергали электрофорезу в редуцирующих условиях (2 % SDS, 5 % 2-меркаптоэтанол) в 13,0 % ПААГ в соответствии с методом Лэммли [13]. Гели окрашивали Кумасси ярко-синим R-250, сканировали и оцифровывали с помощью компьютерной программы Скангель (д-р А.Ф. Равич, ФГБНУ «НИИСХ Юго-Востока»), учитывавшей число пикселей в отдельных фракциях электрофоретического спектра белков. Величину перевариваемости вычисляли как процентное отношение разницы между числом пикселей в контрольном и опытном образце к числу пикселей в контрольном образце. Эксперименты проводили в двух-четырех повторениях.
Статистическую обработку результатов экспериментов осуществляли с использованием пакета программ AGROS, версия 2.09 (д-р С.П. Мартынов, ИОГен РАН).
Результаты исследования и их обсуждение
Анализ перевариваемости запасных белков в системе in vitro у потомства полученных ранее трансгенных растений показал, что некоторые из них отличаются более высокой перевариваемостью, по сравнению с исходной нетрансгенной линией, Ж10. Так, у растений поколения Т3 из семей 94-2-04 и 94-3-04, выращенных в вегетационных сосудах, уровень перевариваемости достигал 85–90 % (рис. 1, табл. 1). При этом в семье 94-2-04 зерновки с более высокой перевариваемостью (83–87 %) отличались модифицированным типом эндосперма, в котором стекловидный слой был представлен в виде тонкого «серпика» (рис. 2), тогда как сегрегант с обычным стекловидным эндоспермом имел низкую перевариваемость (43–55 %, табл. 1). Однако у растений из семьи 94-3-04 высокая перевариваемость наблюдалась у зерновок, как со стекловидным (рис. 1, треки 9–12), так и с мучнистым типом эндосперма (рис. 1, треки 5–8). Следует отметить, что мучнистый тип эндосперма характерен для трансгенных растений кукурузы и сорго с сайленсингом гамма-проламинов [2, 7, 8].
В потомстве трансгенных растений из других семей наблюдалось значительное варьирование перевариваемости запасных белков, связанное, по-видимому, с условиями выращивания. Так, в потомстве растения 94-6 с высокой перевариваемостью кафиринов все четыре изученных растения Т2 (94-6-1, 94-6-2, 94-6-3, 94-6-4), выращенные в условиях климатокамеры, не отличались по уровню перевариваемости от исходной нетрансгенной линии. Однако среди растений из этой семьи, выращенных на опытной грядке, обнаружились растения с почти полной перевариваемостью запасных белков, которая обычно наблюдается у пшеницы или ржи, но не у сорго (рис. 3, треки 10, 12). Данный факт заслуживает дальнейшего детального исследования. Учитывая, что зерновки у таких растений имели обычную текстуру эндосперма с выраженным стекловидным слоем, то данный генотип представляет высокую ценность для возможного использования в селекции. У растений из другой трансгенной семьи, 94-2-04 (поколение Т4), выращенных в условиях опытной грядки, уровень перевариваемости также значимо превышал показатели исходной нетрансгенной линии Желтозерное-10 (табл. 2), что свидетельствует о проявлении данного признака не только в искусственных тепличных условиях, но и в естественных условиях внешней среды.
Таблица 1
Перевариваемость in vitro белков эндосперма трансгенных растений сорго (семьи Т3)
|
Образец |
Тип эндосперма |
Перевариваемость, %1 |
|
Желтозерное-10, исходная линия |
Стекловидный |
52,3 a |
|
Т3 94-3-04-1 |
Мучнистый |
83,1 cd |
|
Модифицированный |
90,3 d |
|
|
Т3 94-02-4-2 |
Мучнистый |
85,2 d |
|
Т3 94-02-4-3 |
Стекловидный |
49,3 a |
|
НСР05 |
10,9 |
|
|
F |
34,105* |
Примечание. 1 Данные, обозначенные разными буквами, значимо отличаются при p < 0,05 в соответствии с Тестом множественных сравнений Дункана; *p < 0,05.
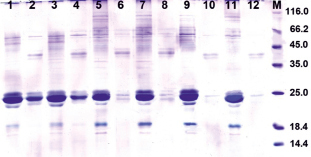
Рис. 1. Перевариваемость кафиринов в системе in vitro у разных зерновок растения Т3 94-3-04-1. 1–4 – исходная нетрансгенная линия Желтозерное-10 со стекловидным эндоспермом; 5–8 – зерновки с мучнистым эндоспермом; 9–12 – зерновки со стекловидным эндоспермом; 1, 3, 5, 7, 9, 11 – контроль без обработки пепсином; 2, 4, 6, 8, 10, 12 – после обработки пепсином; М – маркеры молекулярной массы
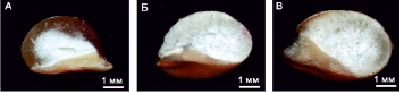
Рис. 2. Поперечные срезы зерновок трансгенных растений сорго. А – зерновка с нормально развитым слоем стекловидного эндосперма, Б – зерновка с мучнистым эндоспермом, В – зерновка с модифицированным типом эндосперма, в котором стекловидный слой представлен в виде «серпика»
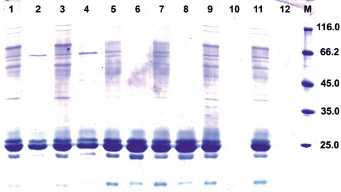
Рис. 3. SDS-электрофорез белков из зерновок трансгенных растений из семьи 94-6 (поколение Т3), выращенных в опытной грядке, до (1, 3, 5, 7, 9, 11) и после обработки пепсином (2, 4, 6, 8, 10, 12). 1–4 – исходная нетрансгенная линия Желтозерное 10, 5–8 – растение из семьи T3 (№ 217-4) с низкой перевариваемостью белков эндосперма, 9–12 – растение из семьи T3 (№ 217-3) с почти 100 % перевариваемостью белков эндосперма; М – маркеры молекулярной массы
Таблица 2
Перевариваемость в условиях in vitro запасных белков трансгенных растений сорго (поколение Т4)
|
Образец |
Условия вегетации |
Перевариваемость, % |
|
|
x ± sx 1 |
Lim |
||
|
Ж10 2 |
Опытная грядка |
53,3 ± 4,4 |
40,0–65,0 |
|
94-2-04-2 |
Опытная грядка |
73,2 ± 3,2 ** |
64,9–81,6 |
|
94-2-11-3 |
Вегетационные сосуды, открытый воздух |
75,0 ± 4,1 ** |
63,5–86,5 |
|
94-2-11-3 |
Вегетационные сосуды, климатокамера |
67,7 ± 8,4 |
47,4–81,8 |
Примечание. 1 sx – ошибка средней; 2 исходная нетрансгенная линия Желтозерное 10; **значимо отличается от исходной линии при p < 0,01, в соответствии с t-тестом Стьюдента.
Заключение
Высокая перевариваемость кафиринов, выявленная нами ранее у трансгенных растений сорго, несущих генетическую конструкцию для индукции сайленсинга гена гамма-кафирина [9], наследуется в их потомстве и проявляется как у растений, выращиваемых в искусственных тепличных условиях, так и в естественных условиях внешней среды. Эндосперм у трансгенных растений с высокой перевариваемостью кафиринов может быть как мучнистым, так и стекловидным, имеющим более высокую ценность в селекции. Данный факт свидетельствует о возможности получения агрономически ценных линий сорго с высокой перевариваемостью кафиринов и твердым эндоспермом.
Библиографическая ссылка
Эльконин Л.А., Итальянская Ю.В. ПЕРЕВАРИВАЕМОСТЬ IN VITRO ЗАПАСНЫХ БЕЛКОВ ЭНДОСПЕРМА ТРАНСГЕННЫХ РАСТЕНИЙ СОРГО, НЕСУЩИХ ГЕНЕТИЧЕСКУЮ КОНСТРУКЦИЮ ДЛЯ САЙЛЕНСИНГА ГЕНА ГАММА-КАФИРИНА // Успехи современного естествознания. 2017. № 12. С. 96-100;URL: https://natural-sciences.ru/en/article/view?id=36612 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/use.36612