


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
IRRIGATION INFLUENCE ON ENTOMOLOGICAL COMPLEX FORMATION OF AGROLANDSCAPES IN LOWER VOLGA AREA

Сельскохозяйственное производство Нижнего Поволжья определяется достаточными земельными и водными ресурсами, потенциально плодородными почвами. Сравнительно высокая теплообеспеченность (t > 10 °C = 2600–3700 °С) и продолжительный безморозный период (150–170 дней) позволяют выращивать здесь не только зерновые колосовые культуры, но и кукурузу на зерно и силос, теплолюбивые овощи (томаты, перец, баклажаны и др.), рис, бахчевые. Значительные площади (до 50 %) заняты кормовыми культурами.
Однако четко выраженная континентальность и недостаточное количество атмосферных осадков (400–180 мм в год при испаряемости 800–1200 мм) приводят к тому, что использование поступающей фотосинтетически активной радиации (18–21 х 109 кДж/га) в условиях естественного увлажнения остается очень низким (0,5–0,1 % и ниже при теоретически возможном 5 %). Иными словами, фактическая продуктивность агробиоценозов в 10–50 раз меньше потенциальной. Реализовать ее можно только при дополнительном увлажнении за счет орошения, которое выступает гарантом устойчивости сельскохозяйственного производства в регионе и в определенной степени способствует стабилизации аграрной экономики.
Однако на орошаемых землях, по сравнению с неорошаемыми массивами, наряду с ростом продуктивности сельскохозяйственных культур возрастает и вредоносность фитофагов, неблагоприятное фитосанитарное состояние отмечается в настоящее время на 77 % сельскохозяйственных угодий, на 70 % засеваемых кормовых угодий и 82 % многолетних насаждений [1–4].
Широкое развитие орошения на юге России обусловливает необходимость всесторонней научно-практической оценки его влияния на формирование энтомокомплексов, изучение особенностей развития насекомых, повреждающих сельскохозяйственные культуры, и в конечном итоге разработки системы защитных мероприятий на орошаемых землях.
В связи с вышеизложенным целью наших исследований являлось изучение влияния орошения на формирование состава и структуры энтомокомплексов хортобионтов и герпетобионтов. В задачу исследований входило изучение видового состава насекомых, определение динамики численности и структуры доминирования комплексов жужелиц в орошаемых и неорошаемых агроландшафтах Волго-Донского междуречья.
Материалы и методы исследования
Исследования проведены в 1987–2017 гг. в орошаемых и неорошаемых агроценозах ФГУП «Орошаемое», расположенного в 20 км западнее г. Волгограда в границах южной части Волго-Донского междуречья. Учеты насекомых-хортобионтов проводили еженедельно в течение вегетационного сезона методом кошения энтомологическим сачком по общепринятой методике Г.Е. Осмоловского с пересчетом на количество экземпляров на 1 м2, герпетобионтов – методом сбора ловушками Барбера [5, 6].
Результаты исследования и их обсуждение
Как показывают наши исследования по определению микроклиматических параметров в агроценозах полевых культур, разница температур на орошаемой и неорошаемой люцерне может достигать 7–8 °С сразу после полива или в среднем за вегетацию до 4–5 °С. При дождевании температура воздуха более интенсивно снижается в течение трех-четырех дней и менее – в продолжение полутора недель. При этом значительно повышается относительная влажность воздуха как внутри травостоя (до 20 %), так и над всем орошаемым участком. Кроме того, на орошении отмечено снижение суточной амплитуды температур. Аналогичные результаты получены в опытах и с другими культурами, причем параметры микроклимата на пропашных (кукуруза, соя) в орошаемых условиях изменяются еще более резко. Указанные следствия влияния орошения, наряду с увеличением влажности почвы, приводят к значительно большей мезофитности микроклимата в агроценозе.
В орошаемых агроценозах отмечено формирование преимущественно полидоминантных энтомологических сообществ со значительным видовым разнообразием. Так, в орошаемом агроценозе люцерны из 254 зарегистрированных видов к доминантам и субдоминантам относится 10–15, тогда как на неорошаемых посевах из 159 зарегистрированных видов преобладают 6–8. В орошаемом агроценозе кукурузы зарегистрировано 86 видов насекомых, из которых 12 включены в доминирующую и субдоминирующую группы, в неорошаемых агроценозах – соответственно 64 и 9 видов. А в целом, по нашим данным, состав энтомофауны орошаемых агроландшафтов Нижнего Поволжья насчитывает 1712 видов, из которых 1432 вида отмечены в агроценозах и 1579 видов – в сопутствующих зональных и интразональных стациях. Видовой состав неорошаемых агроценозов значительно беднее и насчитывает только 982 вида. Анализ структуры энтомокомплексов показал, что в орошаемом агроценозе люцерны из 254 зарегистрированных видов к доминантам и субдоминантам относится 10–15, тогда как на неорошаемых посевах из 159 зарегистрированных видов преобладают 6–8.
В растениях, обеспеченных влагой, изменяется концентрация клеточного сока и сосущая сила листьев, что имеет важное значение для насекомых с колюще-сосущим ротовым аппаратом. Так, согласно данным, представленным на рис. 1, численность люцернового клопа на орошении возросла в 4,6 раза, тли – в 3,3, цикад – в 3,7 раза. Орошение люцерны также благоприятно сказывается и на заселении посевов мезофильными видами. Так, численность люцерновой толстоножки (брухофагуса) на вариантах с орошением по сравнению с неорошаемыми участками увеличилась в 2,7 раза. В результате поврежденность семян брухофагусом возросла в 2,5 раза, а суммарные потери от семеедов – в 2,4 раза.
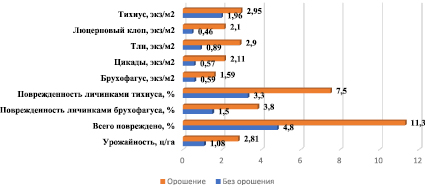
Численность и вредоносность фитофагов на широкорядных посевах 3-го года жизни орошаемой и неорошаемой семенной люцерны (ФГУП «Орошаемое», 1987–2017 гг.)
Орошаемые посевы являются более благоприятными для хищных и паразитических насекомых. При проведении сравнительных учетов численности энтомофауны семенной люцерны установлено, что на орошении численность полезных насекомых оказалась более чем в 7 раз выше, чем на богаре: соответственно 2,51 экз/м2 и 0,35 экз/м2. Численность кокцинеллид возрастает в 6 раз, паразитических перепончатокрылых – в 9 раз, хищных клопов – в 13 раз, златоглазок – в 16 раз. В результате на орошаемых посевах число фитофагов, приходящихся на 1 экземпляр фитофага, более чем в 3 раза ниже, чем в неорошаемых условиях. Это позволяет говорить о том, что на орошаемых посевах создаются более благоприятные условия для естественной регуляции численности вредителей.
Установлено, что под воздействием орошения происходит значительное увеличение обилия энтомофагов напочвенного яруса в агроценозах всех основных кормовых культур (таблица). Данные учетов жужелиц (сем. Carabidae) почвенными ловушками показывают, что на различных культурах сравнительная численность этой группы энтомокомплекса в условиях орошения в 2,5–11,5 раз выше, чем на неорошаемых посевах.
Сравнительное обилие комплексов жужелиц орошаемых и неорошаемых агроценозов ведущих сельскохозяйственных культур по данным учетов в ловушках Барбера (ФГУП «Орошаемое», в среднем за 1987–2017 гг.)
|
Культура |
Орошение, экз./10 л.-с. |
Без орошения, экз./10 л.-с. |
||||||||||||
|
апрель |
май |
июнь |
июль |
август |
сентябрь |
в среднем за сезон |
апрель |
май |
июнь |
июль |
август |
сентябрь |
в среднем за сезон |
|
|
Люцерна |
62,38 |
30,15 |
17,95 |
57,90 |
98,16 |
10,22 |
46,13 |
6,53 |
3,19 |
1,59 |
6,12 |
10,36 |
1,08 |
4,81 |
|
Клевер |
61,02 |
29,15 |
15,84 |
56,01 |
94,20 |
9,84 |
44,34 |
4,87 |
2,75 |
1,34 |
4,56 |
8,85 |
0,75 |
3,85 |
|
Многокомпонентные смеси |
59,16 |
40,89 |
27,87 |
16,74 |
87,15 |
8,27 |
40,01 |
7,31 |
5,13 |
3,53 |
2,17 |
10,35 |
1,17 |
4,94 |
|
Озимая пшеница |
17,25 |
21,10 |
25,97 |
20,01 |
– |
– |
21,08 |
7,09 |
8,54 |
10,62 |
8,24 |
– |
– |
8,62 |
|
Кукуруза |
– |
1,90 |
2,88 |
4,41 |
32,79 |
12,99 |
10,99 |
– |
0,38 |
0,54 |
0,84 |
5,90 |
2,31 |
1,99 |
|
Картофель |
– |
14,79 |
15,88 |
23,11 |
59,58 |
– |
26,01 |
– |
1,29 |
1,41 |
2,48 |
5,31 |
– |
2,62 |
|
НСР05 |
1,44 |
0,09 |
||||||||||||
Увеличение суммарного обилия группировки жужелиц орошаемых агроценозов по сравнению с неорошаемыми происходит в первую очередь за счет увеличения численности на посевах таких видов, как Poecilus cupreus, Pseudoophonus rufipes, Bembidion properans, Clivina fossor, Poecilus nitens, Harpalus distinguendus.
Многолетние исследования, проведенные в орошаемых и неорошаемых ландшафтах Нижнего Поволжья, подтверждают имеющиеся литературные данные о том, что орошение является одним из наиболее мощных антропогенных факторов, воздействующих на экологические условия в агроценозах. На поливных землях, в отличие от неорошаемых, создаются условия, оказывающие влияние как на растения, так и на все живые организмы и, как следствие этого, на распространение вредителей и формирование энтомокомплекса полевых культур [7].
В первую очередь это связано со значительным изменением параметров микроклимата посевов и почвы. Внутри травостоя орошаемых полевых культур складывается микроклимат, характерный скорее для более северных и увлажненных районов России, чем для юго-востока. Естественно, что насекомые в соответствии со своим экологическим стандартом формируют на орошаемых массивах специфичный агробиоценоз, в котором преобладают мезофилы, а плотность популяции ксерофилов значительно снижается.
Отмеченное сглаживание экстремумов температуры и относительной влажности воздуха на орошаемых землях способствует улучшению микроклиматических условий и комфортности обитания насекомых. Следовательно, орошение в аридных регионах следует отнести к одному из основных факторов оптимизации условий жизнедеятельности биотического компонента агроэкосистем и, соответственно, сохранения, а отчасти и расширения его биоразнообразия за счет возрастания численности и видового разнообразия мезофилов и, в меньшей степени, гигрофилов.
В качестве яркого примера приуроченности вида к определенным экологическим условиям являются клубеньковые долгоносики. Так, вид Sitona lineatus L. встречался на орошаемых участках люцерны, а на вариантах без орошения он уступал более сухолюбивому виду S. crinitus Hbst., который занял в наших сборах 100 % всех клубеньковых долгоносиков.
Изменяющаяся экологическая обстановка, с одной стороны, непосредственно влияет на развитие и численность вредителей, с другой – обуславливает выращивание качественно новых растений, что отражается на взаимоотношениях между вредителем и растением, а также на выживаемости, плодовитости, продолжительности жизни отдельных фаз и других биологических особенностях популяции.
Изменение микроклиматических параметров почвы и приземного слоя воздуха приводит также к изменениям фенологии как кормового растения, так и фитофагов. Установлено, что на орошаемых участках прохождение фенологических фаз увеличивается. Так, развитие люцерны на орошении до фазы созревания бобов продолжается на 18–20 дней больше, чем на неорошаемых участках, при этом фенологический цикл развития вредителей на орошении сохраняет свою синхронизацию с фенологией кормового растения, удлиняясь также в среднем от 10 до 20–25 дней. В связи с этим потенциальная вредоносность всех изучавшихся видов вредителей на орошении возрастает. Это объясняется как увеличением периода активного питания личинок и имаго люцернового клопа и личинок фитономуса, так и растянутостью фазы формирования и молочной спелости семян, к которой приурочена откладка яиц самками желтого тихиуса и люцерновой толстоножки (при возделывании люцерны на семена).
Вместе с тем хотелось бы отметить, что создание наиболее благоприятных для развития растений микроклиматических условий на орошаемых землях положительно влияет на ход морфологических, органообразовательных, физиологических и биохимических процессов, способствуя резкому повышению продуктивности растений. При этом генеративные органы формируются более интенсивно. Усиление репродуктивной функции, высокие компенсаторные свойства растений, большая кустистость ведет к увеличению урожайности. И, таким образом, несмотря на рост вредоносности семеедов в 2,4 раза, урожайность семян на орошении также увеличивается, прибавка составляет 161,1 %.
Доминирующие виды жужелиц на орошаемых посевах – Poecilus cupreus, Pseudoophonus rufipes, Bembidion properans, Clivina fossor, Poecilus nitens, Harpalus distinguendus – указаны в качестве активных хищников многих опасных вредителей основных кормовых культур [7–9].
И.Х. Шарова с соавторами [9] отмечает, что в аридных зонах ландшафтное распределение жужелиц отличается неравномерностью. Наиболее высокая плотность видов этого семейства отмечается в мезофитных биотопах, располагающихся в понижениях рельефа. В нашем случае увеличение мезофитности микроклимата агроценоза под влиянием орошения закономерно привело к увеличению обилия населяющих его жужелиц.
Кроме этого, на орошаемых участках значительно возрастает общее число животного населения агроценозов, многие элементы которого создают кормовую базу для хищных жужелиц. Так, в частности, здесь увеличивается обилие злаковых тлей и пшеничного трипса, на возможность питания которыми жужелиц указывает ряд авторов [9]. В связи с этим общее увеличение численности жужелиц при орошении, кроме благоприятного микроклимата, обусловлено также и обилием жертв.
Наряду с увеличением видового разнообразия энтомокомплексов, в орошаемых условиях существенно меняется их трофическая структура. При проведении сравнительных учетов численности энтомофауны установлено, что на орошении относительная численность хищных и паразитических видов насекомых оказалась более чем в 7 раз выше, чем на богаре. Соотношение общей численности энтомофагов и фитофагов составляет здесь 1:1,9, тогда как в условиях богары – 1:6,1.
Выводы
Результаты исследований показывают, что одним из основных факторов оптимизации условий жизнедеятельности биотического компонента агроэкосистем является орошение, способствующее сохранению, а отчасти и расширению его биоразнообразия за счет возрастания численности и видового разнообразия мезофилов и гигрофилов.
Установлено, что изменение микроклиматических параметров почвы и приземного слоя воздуха приводит к изменениям фенологии как кормового растения, так и фитофагов. Так, на орошаемых участках прохождение фенологических фаз увеличивается. Например, на орошаемых посевах люцерны отмечено удлинение развития растений на 18–20 дней от весеннего отрастания до фазы созревания бобов по сравнению с неорошаемыми участками, при этом фенологический цикл развития вредителей на орошении сохраняет свою синхронизацию с фенологией кормового растения, удлиняясь также в среднем от 10 до 20–25 дней.
Создание наиболее благоприятных для развития растений микроклиматических условий на орошаемых землях положительно влияет на ход морфологических, органообразовательных, физиологических и биохимических процессов, способствуя резкому повышению продуктивности растений. Усиление репродуктивной функции, высокие компенсаторные свойства растений, большая кустистость ведут к увеличению урожайности. И, таким образом, несмотря на рост вредоносности семеедов в 2,4 раза, урожайность семян на орошении также увеличивается, прибавка составляет 161,1 %.
Результаты исследований показывают, что в орошаемых условиях отмечено формирование преимущественно полидоминантных энтомологических сообществ со значительным видовым разнообразием. Состав энтомофауны орошаемых агроландшафтов Нижнего Поволжья насчитывает 1712 видов, из которых 1432 вида отмечены в агроценозах и 1579 видов – в сопутствующих зональных и интразональных стациях. Видовой состав неорошаемых агроценозов значительно беднее и насчитывает только 982 вида.
На орошаемых участках отмечено увеличение суммарного обилия группировки жужелиц – активных хищников многих опасных видов фитофагов полевых культур. Увеличение численности жужелиц на орошении объясняется большей мезофитностью микроклимата в напочвенном ярусе стеблестоя, появлением дополнительных экологических ниш, что ведет как к увеличению видового разнообразия, так и к увеличению общей численности комплекса, а также увеличению потенциальной кормовой базы по сравнению с неорошаемым агроценозом, где общее обилие животного населения существенно меньше.
Таким образом, в условиях орошения формируются более стабильные сообщества насекомых за счет увеличения их разнообразия, с одной стороны, и за счет оптимизации их структуры – с другой.
Библиографическая ссылка
Комарова О.П. ВЛИЯНИЕ ОРОШЕНИЯ НА ФОРМИРОВАНИЕ ЭНТОМОКОМПЛЕКСОВ АГРОЛАНДШАФТОВ В НИЖНЕМ ПОВОЛЖЬЕ // Успехи современного естествознания. 2018. № 3. С. 58-64;URL: https://natural-sciences.ru/en/article/view?id=36703 (дата обращения: 01.07.2026).
DOI: https://doi.org/10.17513/use.36703