


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
SEASONAL AND ANNUAL DYNAMICS OF THE GREEN MASS STOCKS IN KOIBELSKAYA STEPPE UNDER CLIMATE CHANGE CONDITIONS

Минусинская котловина – одна из нескольких межгорных впадин, по районированию относится к Южно-Сибирской физико-географической области. Левобережная часть Енисея юга Минусинской котловины известна в литературе как «Койбальская степь» и в административном отношении входит в состав Хакасской автономной области.
В современном рельефе впадины сохранились древние структуры средне-верхнепалеозойского возраста в виде холмисто-грядовых форм рельефа с абсолютными высотами 300–500 м. При моноклинальном залегании пород они приобретают облик куэстовых гряд, холмисто-увалистых форм, которые обуславливают специфику степей Центральной Азии [1]. Почвенный покров в междуречье Абакана и Енисея представлен обыкновенными и южными черноземами и каштановыми почвами. Настоящие степи котловины создают фон степной растительности, которые развиваются на плакорах и склоновых поверхностях. Ведущую роль в сложении сообществ настоящих степей играют дерновинные злаки: ковыль Крылова (Stipa krylovii), овсец пустынный (Helictotrichon desertorum), типчак (Festuca valesiaca), тонконог гребенчатый (Koeleria cristata). Рыхлокустовые злаки: змеевка оттопыренная (Cleistogenes squarrosa), мятлик оттянутый (Poa botryoides). Осоки стоповидная и твердоватая (Carex pediformis, c. duriuscula). Из полукустарничков – полынь холодная (Artemissia frigida), из кустарников – карагана карликовая (Caragana pygmaea) [2].
На склонах южной экспозиции верхних частей и гребневидных вершинах сопок развиты сообщества каменистых степей, образующих группировки настоящих степей. В сложении этих сообществ ведущую роль играют нитрофитные виды: тимьян азиатский (Thymus asiaticus), смолевка енисейская (Silene jenisseensis). По северным склонам сообщества настоящей степи отличает большая видовая насыщенность: ковыль красноватый (Stipa rubens), прострел желтеющий (Pulsatilla turczaninovii), подмаренник настоящий (Galium verum), скабиоза желтая (Scabiosa ochroleuca), володушка козелецелистная (Bupleurum scorzonerifolium). Одним из приспособлений степных растений к неблагоприятным экологическим условиям являются виды с розеточной и полурозеточной формой роста, для Койбальской степи составляют до 38 %. Например, среди степных растений Забайкалья широко распространена розеточная и полурозеточная форма роста, которая представлена разнотравьем (виды Potentilla, Chamaerhodos) и составляет до 80 % [3].
Цель работы: выявление основных закономерностей сезонной и погодичной динамики запасов зеленой массы в фациях полигона-трансекта Койбальской степи в условиях современного климата.
Материалы и методы исследования
Климат котловины резко континентальный и характеризуется большими годовыми и суточными амплитудами колебаний температуры. Наиболее последовательным и выраженным элементом изменения климата является температура воздуха, при этом важным показателем состояния атмосферы, оказывающим наряду с температурой воздуха влияние на трансформацию растительного покрова, являются осадки. Большая часть осадков приходится на летний период, а наименьшее количество осадков выпадает зимой. В среднем для рассматриваемой территории годовая сумма атмосферных осадков за многолетний период с 2000 по 2017 г., по данным метеостанции «Бея», составляет 469 мм. Максимальное количество осадков 618 мм отмечено в 2003 г., минимальное количество осадков 313 мм в 2011 г. Средняя температура воздуха в январе составляет минус 18–19 °, а в июле плюс 19 °. Средняя годовая температура воздуха за период наблюдений превысила среднюю многолетнюю годовую величину на 1,6 °С, достигнув максимального своего значения 4,3 °С в 2007 г. Самая минимальная среднегодовая температура 0,6 °С отмечена в 2010 г. (рис. 1).
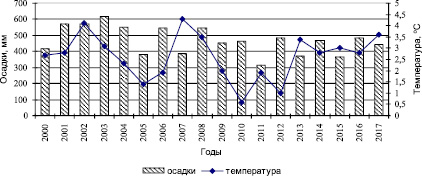
Рис. 1. Погодичная динамика атмосферных осадков и температуры воздуха Койбальской степи (по данным метеостанции «Бея»)
Многолетняя динамика максимальных запасов зеленой массы (г/м2) в фациях полигона-трансекта за период 2000–2017 гг. (абсолютно сухой вес)
|
Годы |
Фации полигон-трансекта |
|||
|
Мелкодерновинно-злаково-ковыльная с караганой (ф. I) |
Петрофитно-разнотравно-ковыльно-типчаковая (ф. II) |
Осоково-овсецово-ковыльная с караганой (ф. III) |
Разнотравно-осоково-овсецово-ковыльная (ф. IV) |
|
|
2000 |
219 |
178 |
205 |
190 |
|
2001 |
183 |
197 |
199 |
183 |
|
2002 |
172 |
141 |
134 |
126 |
|
2003 |
258 |
305 |
264 |
223 |
|
2004 |
275 |
376 |
259 |
142 |
|
2005 |
96 |
94 |
102 |
110 |
|
2006 |
299 |
178 |
170 |
161 |
|
2007 |
230 |
261 |
265 |
113 |
|
2008 |
188 |
224 |
145 |
189 |
|
2009 |
224 |
253 |
194 |
194 |
|
2010 |
261 |
288 |
243 |
242 |
|
2011 |
284 |
296 |
288 |
198 |
|
2012 |
309 |
304 |
322 |
155 |
|
2013 |
182 |
249 |
218 |
182 |
|
2014 |
216 |
217 |
365 |
146 |
|
2015 |
219 |
160 |
165 |
111 |
|
2016 |
190 |
138 |
192 |
119 |
|
2017 |
134 |
98 |
109 |
110 |
Для оценки биологической продуктивности сообществ используются данные общего количества (запаса) растительного вещества надземной части травостоя (зеленая масса). Определение продуктивности исследуемых экосистем проводилось общепринятыми методами [4, 5]. При определении надземной зеленой массы и выявлении особенностей динамики нарастания надземной массы использовали метод «укосных площадок», для определения надземной массы с точностью ±15 % достаточно 3–5 повторностей с площадок по 0,25 м2. Образцы зеленой массы, высушенные до абсолютно сухого состояния, взвешивались на электрических весах (ВЛТК-500). Все результаты записывались в «весовую тетрадь». Полученные данные статистически обработаны и использованы для построения «пространственно-временных моделей», как для сезонной, так и многолетней динамики запасов зеленой массы на полигон-трансекте.
Участок стационарных работ – Новониколаевский полигон-трансект, который пересекает несколько пространственно сопряженных фаций с мелкогрядовыми и плоскоравнинными формами рельефа. Основные фации полигона-трансекта Койбальской степи: ф. I – мелкодерновинно-злаково-ковыльная с караганой степь склона южной экспозиции (т. 7); ф. II – петрофитно-разнотравно-ковыльно-типчаковая степь на вершинной поверхности (т. 15); ф. III – осоково-овсецово-ковыльная с караганой степь на выровненной поверхности (т. 36); ф. IV – разнотравно-осоково-овсецово-ковыльная степь, денудационный склон северной экспозиции (т. 42).
Результаты исследования и их обсуждение
Закономерности пространственного распределения максимальных запасов зеленой массы удобнее рассмотреть по элементам рельефа полигона-трансекта. Для выровненных поверхностей (ф. III) с растительным покровом – осоково-овсецово-ковыльным с караганой, за годы наблюдений зеленая масса изменялась от 102 (2005 г., сухой), до 365 г/м2 (2014 г., влажный) (таблица). Во влажные годы величина зеленой массы увеличивается в 3,5 раза. Для выпуклых вершин (ф. II) с петрофитно-разнотравно-ковыльно-типчаковым сообществом выявлена прямая зависимость надземной массы от количества осадков, которые варьируют от 94 (2005 г., сухой) до 376 г/м2 (2004 г., влажный), запасы во влажный год были выше в 4 раза. В растительном покрове южных склонов (Ф. I), где общий фон растительности создают мелкодерновинно-злаково-ковыльные сообщества с караганой карликовой, максимальные запасы зеленой массы составляют от 96 (2005 г.) до 309 г/м2 (2012 г., влажный). Интересной особенностью природы северных крутых склонов, где поверхности характеризуются горным обликом рельефа, в этих условиях растительные сообщества отличаются большим видовым разнообразием: красочностью аспектов и большой продуктивностью. Величина зеленой массы в разнотравно-осоково-овсецово-ковыльном сообществе (Ф. IV) составила 45 % от максимальной. Максимальная величина составляет 242 г/м2 (2010 г.).
Принимая во внимание среднемаксимальные показатели (по мере их роста) зеленой массы за все (с 2000 по 2017) годы исследований, изученные фации можно расположить в такой ряд: II > I > III > IV.
На построенных моделях по запасам фитомассы топохроноизоплеты наглядно показывают флуктуации зеленой массы по годам и фациям, при одинаковом поступлении тепла и влаги. Отображение пространственно-временных изменений запасов массы в сопряженном ряду фаций полигона-трансекта представляет собой стационарную модель Койбальской степи (рис. 2).
Стационарные наблюдения в степных фациях показали значительные колебания зеленой массы по годам. Нарастание фитомассы – процесс ритмичный, ежегодно повторяющийся в общих своих чертах. В зависимости от погодной обстановки и внутренних ритмов развития в больших пределах меняется от года к году. Сравнивая два смежных года (2004 и 2005), видим, что первый характеризуется повышенной величиной запасов зеленой массы во влажный год на ф. II, а второй – пониженной величиной в сухом годе на всех фациях (рис. 2). Количественно оценить зависимость запасов зеленой массы от погодных условий довольно затруднительно, ибо травостой по-разному реагирует на условия окружающей среды, т.е. имеет свои особые ритмы развития. Причем каждая фация имеет свои характеристики периодичности в изменении фитомассы и свои амплитуды ее колебаний.
Исследования по определению особенностей формирования надземной фитомассы, ее сезонной и погодичной динамики выявили цикличность – сухие периоды с угнетенным состоянием растений – цикл сухих лет (2002, 2005, 2015 и 2017), сменяющиеся влажными сроками с хорошим травостоем – цикл влажных лет (2003, 2004, 2007 и 2014 гг.). Накопление зеленой массы достигается только в конкретный для нее срок, что подтверждается данными других исследователей в степях Хакасии, Тувы и Монголии [6–8].
Наблюдения за сезонной динамикой зеленой массы показывают четкую ритмичность, не только по месяцам, но и в сопряженном ряду фаций полигона-трансекта (рис. 3, а, б).
Высокая амплитуда колебаний климатических факторов, контрастность погодных условий в течение вегетационного сезона приводит к резким колебаниям фитомассы, тем самым определяя особенности функционирования степных сообществ. Средние за четыре года – период сухих лет (2002, 2005, 2015 и 2017 гг.) показывают, что формирование зеленой массы в течение сезона вегетации полностью зависит от метеорологических условий и в первую очередь от осадков (рис. 3, а). Чрезвычайно низкая величина фитомассы (зелени) от 25 до 50 г/м2 в апреле, мае месяце при температуре 4,8 °С, высота травостоя в этот период была выражена слабо. Расчленение травостоя на подъярусы не было заметно. Новые побеги плохо росли, из-за малого количества осадков от 23 до 28 мм, в течение этих двух месяцев. Осадки, выпавшие в июне до 70 мм, стимулировали увеличение зеленой массы в июле месяце до 143 г/м2. Аномальные по метеоусловиям засушливые годы приводят к уменьшению контрастности показателей запасов зеленой массы в разных условиях рельефа с небольшим увеличением на южном склоне (ф. I). Запасы фитомассы на всех фациях в апреле и мае месяце составляли от 28 до 67 г/м2. Максимальная величина однолетней массы травостоя отмечена на (ф. I) в июле (рис. 3, б). Выпавшие в августе осадки стимулировали незначительное отрастание позднелетних видов растений с небольшим увеличением в массе до 115 г/м2 (ф. I, II), в сентябре происходит уменьшение запасов на всех фациях, а на (ф. IV) составляют всего 85 г/м2.
Средние данные за четыре года – период влажных лет (2003, 2004, 2007 и 2014 гг.) показывают, что в формировании зеленой массы прослеживается четкая ритмичность, не только по месяцам, но и в сопряженном ряду фаций полигона-трансекта (рис. 4, а, б).
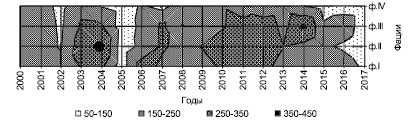
Рис. 2. Пространственно-временная изменчивость запасов зеленой массы на полигон-трансекте Койбальской степи, в многолетнем ряду, г/м2
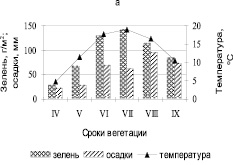
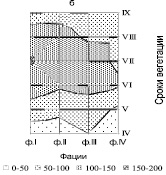
Рис. 3. а) сезонная динамика зеленой массы, осадков и температуры за вегетационный период сухих лет (2002, 2005, 2015 и 2017); б) пространственно-временные колебания зеленой массы за сухой период. Топохроноизоплетами показаны запасы зелени; I – IV – фации
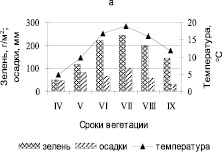

Рис. 4. а) сезонная динамика зеленой массы, осадков и температуры за период влажных лет (2003, 2004, 2007 и 2014 гг.); б) пространственно-временные колебания зеленой массы за период влажных лет. Топохроноизоплетами показаны запасы зелени; I – IV – фации
Нарастание зеленой массы в ранневесенний период (в апреле, мае), выпавшие осадки до 82 мм при температуре до 10 °С оказали существенное влияние на формирование растительного покрова. Май месяц был более влажным, чем апрель, и предшествовал накоплению зеленой массы в июне до 221 г/м2. В июле месяце (ф.I I, III) зеленая масса была выше в 2 раза (290 г/м2), по сравнению с сухим периодом. Максимальная величина отмечена в конце июля, в начале августа – составила 245 г/м2, при температуре в 19 °С и осадках 101 мм. Однако в связи с уменьшением количества осадков в августе и сентябре месяце, при устойчиво высоких температурах воздуха и небольших осадках травостой оставался без значительных изменений. Заканчивая свой позднеосенний вегетационный период в сентябре, октябре с уменьшением осадков до 32 мм и температуры до 12 °С, масса зелени составляет 144 г/м2 (рис. 4, а). Наблюдения показали, что в годы с повышенным количеством осадков в апреле и мае (48–82 мм) способствуют накоплению фитомассы в июне, июле месяце, хотя количество осадков в этих сроках не увязывается с величиной создаваемой зеленой массы. Небольшие осадки, а зеленая масса высокая. Следовательно, основным прямодействующим фактором оказались осадки за апрель – май.
Пространственно-временные колебания зеленой массы в фациях полигона-трансекта позволяют понять картины утраченного в сухие годы и выявить тенденции развития ее во влажные годы в фациях степных сообществ Койбальской степи (рис. 4, б). Максимальные запасы для большинства фаций формируются в июле, этот максимум выражен более четко, по сравнению с фациями сухого периода. Так средние за четыре года запасы зеленой массы в середине июля составляют от максимального укоса до 80 % на ф. I, II и III, исключая ф. IV – разнотравно-осоково-овсецово-ковыльную, склона северной экспозиции с раннелетним ритмом накопления фитомассы (рис. 4, б).
Выводы
1. Величина запасов зеленой массы Койбальской степи в довольно больших пределах колеблется от года к году. Травостой по–разному реагирует на условия окружающей среды, имеет свои особые ритмы развития. Каждая фация имеет свои характеристики периодичности в изменении зеленой массы и свои амплитуды колебаний.
2. Установлены значительные изменения величины зеленой массы от сезонных и пространственно-временных колебаний в разных элементах рельефа полигона-трансекта. Аномальные по метеоусловиям засушливые годы приводят к уменьшению контрастности показателей запасов зеленой массы в разных условиях рельефа с небольшим увеличением на южном склоне (ф. I, рис. 3, б).
3. Выявлены значительные изменения величины зеленой массы в вегетационном периоде и в фациях полигона-трансекта во влажные годы. Максимальные запасы для большинства фаций формируются в июле, этот максимум выражен более четко (ф. II, III, рис 4, б), зеленая масса была выше в 2 раза, по сравнению с сухим периодом.
Библиографическая ссылка
Дубынина С.С. СЕЗОННАЯ И ПОГОДИЧНАЯ ДИНАМИКА ЗАПАСОВ ЗЕЛЕНОЙ МАССЫ КОЙБАЛЬСКОЙ СТЕПИ В УСЛОВИЯХ ИЗМЕНЕНИЯ КЛИМАТА // Успехи современного естествознания. 2018. № 6. С. 65-70;URL: https://natural-sciences.ru/en/article/view?id=36783 (дата обращения: 08.08.2026).
DOI: https://doi.org/10.17513/use.36783