


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
Anatomic-morphological features of the Pinus sylvestris L. mycotrophy in the plantations damaged by the Heterobasidion annosum (Fr.) Bref.

Heterobasidion annosum – возбудитель стволовой гнили древесины, на сегодняшний день он является основным патогеном Pinus sylvestris L. в умеренных широтах. Корневая губка массово распространена в чистых сосновых культурах, созданных во многих областях России, особенно в тех местах, где сосна посажена на площадях, которые длительное время не были заняты лесом, таких как пастбища, старопахотные земли и т.д. [1]. Отмечается тенденция к усилению распространения Heterobasidion annosum (Fr.) Bref. в связи с изменением климата, а следовательно, и возрастание экономического ущерба, наносимого лесному хозяйству [2]. В настоящее время проводятся многочисленные работы по выработке мер противодействия корневой губке. Проводятся исследования вирусов, паразитирующих на Heterobasidion annosum [3]. Исследуются защитные реакции саженцев хвойных, отмечено сходство ответной реакции растения на заражение Heterobasidion annosum с реакцией на заражение микоризным грибом Pisolithus tinctorius [4]. Учитывая тот факт, что сосна обыкновенная является высокомикотрофным растением, не способным к нормальному развитию без микориз, их роль в формировании устойчивости Pinus sylvestris к корневой губке может быть весьма значительной.
У сосны обыкновенной описаны три основных типа метаморфизированных корневых окончаний, образованных в результате взаимодействия гриба и высшего растения: эктотрофные микоризы, эктоэндотрофные микоризы и псевдомикоризы. При благоприятных условиях произрастания на корневых окончаниях сосны формируются эумицетные хальмофаговые эктомикоризы, или эктотрофные микоризы [5]. Эктотрофная микориза образуется главным образом у хвойных и «сережкоцветных» покрытосеменных. Трофические связи симбионтов сводятся к хальмофагии или резорбции солей и других веществ. У хвойных взаимоотношения гриба и высшего растения варьируют от факультативного до облигатного симбиоза, но главным образом сводятся к двустороннему умеренному паразитизму гриба и высшего растения, или аллелопаразитизму [6]. Эндоэктотрофные микоризы отмечались прежде всего в хвойных питомниках и в биотопах с нарушенными условиями обитания. Псевдомикоризы в здоровых лесных условиях встречаются редко и наблюдаются в случаях, когда растение сильно подавлено, например при выращивании сеянцев хвойных в стерильных условиях. Псевдомикоризы являются односторонним паразитизмом слишком вирулентных грибов на высшем растении. При определенных условиях или на определенной стадии в ходе развития ассоциации роль микоризного гриба может меняться. Один и тот же микобионт может быть эндофитом, мутуалистическим симбионтом, сапротрофом или некротрофным паразитом. Грибы, образующие эктомикоризы с одним растением, у другого могут формировать эктоэндомикоризы и даже псевдомикоризы [6].
Исследование особенностей корневого питания Pinus sylvestris в насаждениях, пораженных Heterobasidion annosum, особенно актуально, поскольку может помочь в выработке мер борьбы с корневой губкой. Цель нашего исследования – изучить анатомо-морфологические особенности микотрофности Pinus sylvestris в насаждениях, пораженных Heterobasidion annosum и в здоровых древостоях.
Материалы и методы исследования
Пробные площади (ПП) закладывались в лесных культурах Pinus sylvestris, созданных ручной посадкой на старопахотных землях Клюковенского участкового лесничества Государственного казенного учреждения Брянской области «Навлинское лесничество». Насаждения представляют собой лесные культуры: возраст 44 года, состав 10С, ТЛУ (тип лесорастительных условий) – B2, тип леса – сосняк орляковый. В насаждениях зафиксировано до 10 % деревьев, пораженных Heterobasidion annosum.
Микоризы сосны и других хвойных пород – это однолетние образования, функционирующие в течение одного вегетационного периода, а затем погибающие. Рост корней характеризуется определенным ритмом, развитие микориз очень тесно связано с изменениями активности роста дерева-хозяина и условий среды, окружающей корни. Это обстоятельство имеет особенное значение при выборе сроков отбора материала для исследований [5]. Сбор полевого материала осуществлялся в конце вегетационного периода, причем отбор образцов корневых систем осуществлялся в течение 4…7 дней, что позволило свести к минимуму климатические отличия на ПП и считать их одинаковыми на опытных и контрольных площадях. Отобрано и проанализировано 700 корневых окончаний сосны обыкновенной.
Из отобранных материалов приготавливали микросрезы толщиной 5…10 мкм. Полученные препараты исследованы с использованием микроскопов МБИ-6 и Микмед. Размеры исследуемых элементов микориз определяли при помощи окуляра, оснащенного шкалой Гюйгенса. Разнообразие эктомикориз исследовано с использованием классификации, изложенной в работе И.А. Селиванова [6], основанной на признаках анатомического строения эктомикоризных чехлов, выделяемые при этом способе классификации единицы – подтипы микоризных чехлов. В корневых окончаниях измеряли толщину грибного чехла, толщину паренхимы первичной коры, радиус центрального цилиндра, общую толщину корневого окончания. Статистическая обработка экспериментального материала осуществлялась по схеме малой выборочной совокупности, при этом определялись такие статистические показатели, как средняя арифметическая величина с ошибкой (М ± mx), среднее квадратическое отклонение (s), точность опыта (р). Достоверность различия средних величин оценивалась по t-критерию Стьюдента. Все вычисления осуществлялись с использованием пакета прикладных программ Microsoft Excel на ЭВМ.
Результаты исследования и их обсуждение
Микоризы сосны имеют простую (неразветвленную), вильчатую, дваждывильчатую и сильноразветвленную (коралловидную) формы.
Типичная эндотрофная микориза (или эндомикориза), характеризующаяся отсутствием грибного чехла, сохранением неизменным внешнего вида корневого окончания и наличием обильных грибных структур в клетках корня, у Pinus sylvestris нами не была обнаружена.
В очаге поражения Heterobasidion annosum нами зафиксированы немикоризные корневые окончания, эктотрофная микориза и псевдомикориза (рисунок).
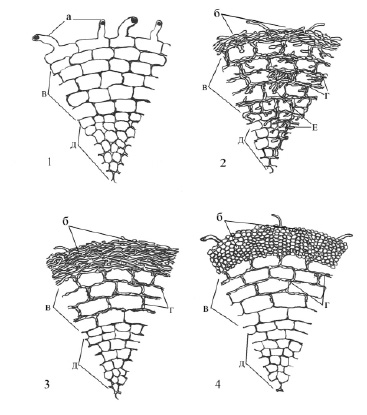
Корневые окончания сосны обыкновенной в насаждениях, поврежденных Heterobasidion annosum (Fr.): 1 – Немикоризное корневое окончание; 2 – псевдомикориза; 3 – эктотрофная микориза с плектенхиматическим строением грибного чехла; 4 – эктотрофная микориза с псевдопренхиматическим строением грибного чехла; а – корневые волоски; б – грибной корневой чехол; в – первичная кора; г – сеть Гартига; д – центральный цилиндр; е – мицелий гриба в клетках корня
На контрольных ПП, заложенных в здоровых насаждениях, псевдомикориза не была обнаружена ни в одном образце.
Микоризность сосны (доля метаморфизированных корневых окончаний, к которым мы причисляем и псевдомикоризу) составила 93–97 %, статистически достоверных отличий на контрольных и опытных ПП не отмечено.
Эктотрофная микориза сосны отличается наличием грибного чехла, развитием сети Гартига, отсутствием внутриклеточной инфекции.
При образовании микориз и формировании грибного чехла корневые волоски редуцируются. Сеть Гартига хорошо развитая, проникает по межклетникам на три-пять слоев клеток первичной коры. Гифы гриба способны внедряться внутрь клеток корня, но очень незначительно и ограничиваются периферическим слоем клеток, богатых дубильными веществами. Внутри этих клеток происходит переваривание мицелия, гифы разрушаются, образовавшиеся гранулы затем уменьшаются и постепенно исчезают. Часть периферических клеток первичной коры (от нескольких клеток до нескольких слоев клеток) обычно представлена таниновыми клетками, которые имеют темно-коричневую, иногда черную окраску, более плоскую форму, часто наблюдается потеря ими тургора. Таниновые клетки встречались нам в микоризах со всеми подтипами чехлов, (микориз с бесструктурными чехлами, не содержащих таниновых клеток, авторами не зафиксировано). Часть периферических слоев клеток первичной коры (как правило, 1 или 2 слоя) бывают представлены таниновыми клетками. В немикоризных корневых окончаниях таниновые клетки не отмечены.
Авторами обнаружены эктотрофные микоризы с различными подтипами грибных чехлов. Микоризы с плектенхиматическим (войлочным) строением грибного чехла, на ПП зафиксированы подтипы: В – поверхность грибного чехла гладкая или покрыта слабой бахромой, С – на поверхности грибного чехла встречаются ризоморфы, D – грибной чехол покрыт щетинками, А – имеет рыхлое, неправильное соединение гифов с корнем. Отмечены микоризы с псевдопаренхиматическими подтипами, среди них: F – имеет гладкую или с одиночными гифами поверхность грибного чехла, H – грибной чехол покрыт многочисленными светлыми, мягкими гифами; G – поверхность грибного чехла покрыта жесткими, тупоконечными гифами. Микоризы с двойными чехлами представлены тремя подтипами: P – наружный слой грибного чехла светлый, плектенхиматического строения, внутренний – темный, псевдопаренхиматический, K – наружный слой гладкий, коричневого цвета, плектенхиматического строения, внутренний – псевдопаренхиматический, другой окраски, О – имеет темный наружный слой и светлый внутренний, оба псевдопаренхиматического строения. Из микориз с чехлом бесструктурного строения отмечен только один подтип – S, характеризующийся гладкой, более или менее однородной поверхностью.
Поскольку анатомо-морфологические показатели, например толщина грибного чехла, зависят от типа микориз, целесообразно исследовать их по основным типам. Среди эктомикориз сосны бесструктурные и двойные чехлы встречались на ПП в незначительном количестве. Наиболее распространены плектенхиматические и псевдопаренхиматические подтипы микориз, они встречаются на всех ПП, анализ анатомо-морфологических особенностей эктомикориз авторы проводили по этим подтипам.
Немикоризные корневые окончания характеризуются наличием корневых волосков, отсутствием грибного чехла и сети Гартига. В паренхиме их первичной коры авторам также не встречались таниновые клетки и клетки с потерей тургора.
Псевдомикоризы сходны по внешнему виду с истинными микоризами, отличить их можно лишь под микроскопом. На поперечном срезе можно видеть значительные отклонения псевдомикориз от эктомикориз. У псевдомикориз меньше, чем у истинной микоризы, развит грибной чехол, клетки растения заражены грибом и часто наполнены гаусториевидными гифами, прокладывающими себе путь от обволакивающего корень грибного чехла. Причем гифы проникают почти в клетки коровой ткани и центрального цилиндра. Заражение корня разветвленными пальцевидными гифами, напоминают картину заражения некоторыми паразитами. В классических псевдомикоризах менее выражена или отсутствует сеть Гартига [6]. Псевдомикоризы с полным отсутствием сети Гартига обнаружены не были, что, наряду с имеющимся грибным чехлом, позволяет отнести их к псевдоэктомикоризам. У корневых окончаний этого типа отсутствуют признаки лизиса (внутриклеточного переваривания) гифов гриба. Это может свидетельствовать о нарушении нормально уравновешенных соотношений между деревом-хозяином и грибом, переход гриба к одностороннему паразитизму.
В ходе исследования анатомо-морфологических параметров корневых окончаний наименьшие размеры грибного чехла (13,6 ± 0,6 мкм) зафиксированы авторами у псевдомикориз. Толщина паренхимы первичной коры псевдомикориз составила 74,9 ± 3,4 мкм, радиус центрального цилиндра – 55,8 ± 2,7 мкм, общая толщина корневого окончания равна 288,6 ± 9,1 мкм.
Толщина грибного чехла эктомикориз плектенхиматических подтипов составила 17,0 ± 0,8 мкм, что на 3,4 мкм больше аналогичного показателя псевдомикориз, различие статистически достоверно (tSt > tтабл при р = 0,01). Общая толщина плектенхиматических микориз равняется 294,2 ± 6,7 мкм, радиус центрального цилиндра – 56,9 ± 1,7 мкм, толщина паренхимы первичной коры – 73,2 ± 2,8 мкм, статистически значимых отличий от псевдомикориз не обнаружено (таблица).
Анатомо-морфологические показатели корневых окончаний Pinus sylvestris в насаждениях, поврежденных Heterobasidion annosum
|
Тип корневого окончания |
Толщина грибного чехла, мкм |
Толщина паренхимы первичной коры, мкм |
Радиус центрального цилиндра, мкм |
Толщина корневого окончания, мкм |
|
Псевдомикоризы |
13,6 ± 0,6 |
74,9 ± 3,4 |
55,8 ± 2,7 |
288,6 ± 9,1 |
|
Эктомикоризы с плектенхиматическими подтипами грибного чехла |
17,0 ± 0,8 |
73,2 ± 2,8 |
56,9 ± 1,7 |
294,2 ± 6,7 |
|
Эктомикоризы с псевдопаренхиматическими подтипами грибного чехла |
24,3 ± 0,7 |
73,2 ± 2,4 |
56,6 ± 1,7 |
308,0 ± 6,7 |
У микориз с псевдопаренхиматическими подтипами грибных чехлов размеры составляют: толщина паренхимы первичной коры – 73,2 ± 2,4 мкм, радиус центрального цилиндра – 56,6 ± 1,7 мкм, толщина грибного чехла – 24,3 ± 0,7 мкм, общая толщина – 308,0 ± 6,7 мкм. Как и у плектенхиматических подтипов, у псевдопаренхиматических микориз значимое отличие (tSt > tтабл при р = 0,001) зафиксировано только по толщине грибного чехла, который почти в два раза превышает толщину аналогичного показателя псевдомикориз (24,3 ± 0,7 мкм).
Микоризы имеют первостепенное значение для растений и деревьев, растущих на целинных лесных почвах и родственных типах почв. Известно, что псевдомикориза может образоваться вследствие заражения растения слабопатогенным грибом [6].
При создании лесных культур на старопахотных землях отсутствие грибов, образующих эктотрофную микоризу, или наличие слабопатогенных грибов, образующих псевдомикоризу, может привести к ослаблению растения, что делает его более восприимчивым к поражению Heterobasidion annosum. Также причиной возникновения псевдомикориз может быть ослабление растения, вызванное развитием корневой губки, что может привести к нарушению нормальных мутуалистических отношений между корешком и мицелием истинного микоризного гриба. В здоровых насаждениях сосны обыкновенной псевдомикориза не обнаружена ни в одном образце.
Для диагностирования характера взаимоотношений между сосной обыкновенной и грибом имело бы большое практическое и научное значение коллекционирование препаратов типичных микориз и псевдомикориз.
Заключение
Развитие у Pinus sylvestris псевдомикориз, в насаждениях, поврежденных Heterobasidion annosum, может свидетельствовать о нарушении нормально уравновешенных соотношений между деревом-хозяином и грибом, переход гриба к одностороннему паразитизму. Изучение микотрофности Pinus sylvestris в насаждениях, поврежденных Heterobasidion annosum, может помочь в выработке мер борьбы с корневой губкой.
Библиографическая ссылка
Адамович И.Ю. Анатомо-морфологические особенности микотрофности Pinus sylvestris L. в насаждениях, поврежденных Heterobasidion annosum (Fr.) Bref. // Успехи современного естествознания. 2018. № 7. С. 26-31;URL: https://natural-sciences.ru/en/article/view?id=36798 (дата обращения: 02.08.2026).