


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
SPECIFIC FEATURES OF FINE ROOT FORMATION AT DIFFERENT STAGES OF REGENERATION OF POSTAGROGENIC ECOSYSTEMS IN SOUTHERN TAIGA ZONE

Многочисленные исследования показывают, что залежные земли служат устойчивым стоком углерода. По данным Ю.И. Баевой и соавторов [1] в почвах различных природно-климатических зон происходит достоверное увеличение запасов органического углерода в ряду пашня – залежь – лес. Большинство работ по изучению постагрогенных ландшафтов в различных природно-климатических зонах посвящено оценке содержания углерода в почвах и растительном покрове, особенностям изменения видового состава растительности. При этом процессам накопления углерода подземными органами растений уделяется недостаточно внимания.
Известно, что от 50 до 70 % углерода почвы происходит от корней и корневых микроорганизмов [2, 3]. В некоторых экосистемах тонкие корни являются основным источником поступления органического вещества в почву. На их производство затрачивается около трети глобальной ежегодной чистой первичной продукции [4], а органическое вещество, поступающее от разложения тонких корней, является более устойчивым по сравнению с органическим веществом от наземных органов растения [5].
Тонкие корни <2 мм играют ключевую роль в регулировании биогеохимических циклов экосистемы. Результаты исследований N. Makita и соавторов [6] показывают, что при оценке корневой биомассы и морфологических особенностей необходима более подробная классификация тонких корней <2,0 мм. Очень тонкие корни диаметром <0,5 мм могут проявлять видоспецифические черты и изменять их потенциал для поглощения питательных веществ и воды в зависимости от глубины.
Таким образом, изучение морфологических признаков тонких корней различных классов диаметра в гетерогенных растительных сообществах является важным для понимания внутрипочвенных процессов депонирования углерода. Особенно актуальны подобные исследования на последовательных стадиях постагрогенной эволюции сельскохозяйственных угодий, так как они позволяют прогнозировать накопление углерода в почвах и оценить процессы восстановления экосистемы в целом.
Цель исследования: провести сравнительный анализ морфологических признаков тонких корней растительных сообществ на последовательных стадиях восстановления постагрогенных экосистем в зоне южной тайги.
Материалы и методы исследования
Для изучения морфологических признаков тонких корней на переходной стадии самовосстановления нарушенной экосистемы в северной окраинной части Амуро-Зейской равнины выбрано три фитоценоза, и заложено три учетных площадки по 50 м2: ненарушенный ценоз – лес (53 °43'49,7"; 127 °04'01,1"), залежь (53 °43'50,1"; 127 °03'53,1") и сенокос (53 °44'33,4"; 127 °05'37,9") (рис. 1).
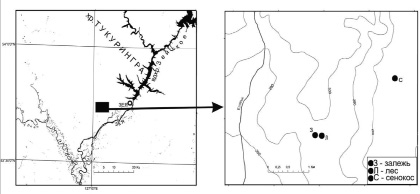
Рис. 1. Схема расположения объектов исследования (З – залежь, Л – лес, С – сенокос)
Климат северной окраинной части Амуро-Зейской равнины характеризуется как умеренно холодный, континентальный с муссонным характером распределения осадков, формируется в результате взаимодействия тихоокеанского муссона и сибирского антициклона. В исследуемом районе средняя многолетняя температура воздуха января минус 28,8 °С, июля 19,7 °С, среднегодовая минус 0,7 °С.
Большую часть территории занимают бурые лесные почвы (буроземы). В целинном состоянии они имеют подстилку мощностью не более 3 см, обладают четкими переходами генетических горизонтов. В стадии пашни почвы характеризуются очень низким содержанием гумуса. Буроземы используются под зерновые, технические, кормовые, плодовые и овощные культуры. Ввиду интенсивного освоения земель в 1950–1970-х гг. большие площади этих почв в Приамурье были вовлечены в пашню, в 1990-е гг. заброшены из-за низкого плодородия и экономического кризиса.
Для установления продуктивности подземной части экосистемы, оценки морфологических признаков тонких корней в августе 2017 г. произведен отбор почвенных образцов в 5-кратной повторности на глубину 0–20 см с помощью металлического цилиндра с диаметром 5 см. На каждой из учетных площадок пробы отбирали в пяти точках (четыре по углам и одна в центре). На сенокосе при определении точки отбора образца выбирали площадку с типичным и ровным травостоем, на площадках «лес» и «залежь» – участки с преобладающей в данном фитоценозе растительностью.
В полевых условиях почвенный образец с корнями разделяли на слои: верхний (0–10 см) и нижний (10–20 см). Тонкие корни (<0,5 мм; 0,5–2 мм) отделяли от почвенных образцов путём промывания проточной водой под высоким давлением на сите с размером ячейки 0,25 мм. В работе анализируется биомасса, определённая после высушивания образцов живых корней до постоянного веса при температуре 40 °С. Под общей биомассой корней в работе понимается масса всех тонких корней диаметром менее 2 мм. Диаметр и длина корней определены на программно-аппаратном комплексе анализа морфологии корневых систем WinRhizoRegular + STD4800 (Regent Instruments, Канада). Удельная длина корней (УДК) – длина единицы сухой массы корня, определялась как отношение длины к массе корней (м/г).
Леса северных районов Амурской области занимают 60–65 % территории области [7]. Наибольшие их площади сосредоточены в Зейском (район исследования) и Селемджинском районах. Территория исследования расположена в подзоне южной тайги. Здесь широко распространены лиственнично-сосново-белоберезовые древостои (50–70 % лиственницы, 10–20 % сосны, 10–50 % березы белой и осины). В южнотаежной подзоне, как и в средней тайге, встречаются рододендроновые, брусничные, багульниковые лиственничники.
Для оценки современного состояния изучаемых фитоценозов на различных стадиях постагрогенной эволюции производили описание лесорастительных условий (древостоя, подроста, подлеска, травянисто-кустарниковой растительности).
Лесной участок (площадь 68 га), выбранный для размещения учетной площадки «лес», является типичным для региона исследований. Древостой в лесу образован березой плосколистной Betula platyphylla Sukacz., лиственницей Гмелина Larix gmelinii (Rupr.) Rupr. и осиной Populus tremula L. В кустарниковом ярусе леса преобладает рододендрон даурский Rhododendron dauricum L., ольховник кустарниковый Duschekia fruticosa (Rupr.) Pouzar. Травянистый покров состоит из майника двулистного Maianthemum bifolium (L.) F.W. Schmidt, брусники Vaccinium vitis-idaea L.
Залежь (площадь 45 га), выведена из сельскохозяйственного оборота 20 лет назад. Растительный покров образован кустарниками: ива Salix sp., малина Комарова Rubus komarovii Nakai, с преобладаем травянистых растений: тысячелистника обыкновенного Achillea millefolium L., осоки, полыни Artemisia sp., мелколепестника Erigeron sp., ястребинки зонтичной Hieracium umbellatum L., клевера люпиновидного Lupinaster pentaphyllus Moench и других видов.
Сенокос (площадь 58 га). На сенокосе произрастают пижма Tanacetum sp., одуванчик Taraxacum sp., хлопушка обыкновенная Oberna behen (L.) Ikonn., горошек Vicia sp., герань сибирская Geranium sibiricum L., вейник Calamagrostis sp., спорыш Polygonum sp., репешок мелкобороздчатый Agrimonia pilosa Ledeb., гравилат алеппский Geum aleppicum Jacq., лапчатка Potentilla sp.
Результаты лабораторных и полевых исследований обработаны методами математической статистики в программе R версия 3.3.2. Статистические различия между изучаемыми параметрами установлены при помощи дисперсионного анализа (ANOVA). Проверку гипотез на статистическую достоверность различий и значимость рассчитанных статистик осуществляли при 5 %-ном уровне значимости.
Результаты исследования и их обсуждение
Общая биомасса тонких корней <2,0 мм в слое почвы 0–20 см максимальна на залежи (402 г/м2), что значимо выше по сравнению с сенокосом (259 г/м2) и лесом (295 г/м2) (p<0,001). Большая часть общей биомассы сконцентрирована в верхнем слое почвы 0–10 см (рис. 2).
Дифференциация корней <2 мм на более тонкие фракции показала, что в верхнем слое почв всех исследуемых сообществ преобладают корни <0,5 мм. На стадии сельскохозяйственного использования земель (сенокос) их доля составила 73 % от общей биомассы тонких корней, на залежи – 89 %, в естественной экосистеме (лес) – 61 %. В почвенном слое 10–20 см в лесу доля корней <0,5 мм снижается, тогда как на залежи и сенокосе возрастает.
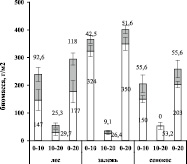
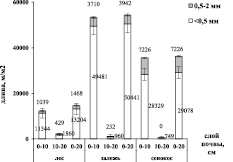
Рис. 2. Биомасса и длина тонких корней по классам диаметра (<0,5 мм и 0,5–2,0 мм) и глубине почвенного слоя (0–10 см, 10–20 см, 0–20 см), планки погрешностей отражают ошибки средних величин (n = 5)
На агроценозе (сенокос) длина корней <0,5 мм в верхнем слое почвы в 2,5 раза больше, чем в лесной экосистеме (p < 0,001), в то время как их биомасса значимо не различалась (p = 0,999). F. Bredoire и соавторы [8] отмечают, что длина тонких корней, как правило, больше на лугах, в то время как общая масса тонких корней, как правило, больше в лесах. Авторы предполагают, что это происходит за счет разницы в диаметре и/или плотности корневых тканей деревьев и трав.
Процесс трансформации нарушенных экосистем характеризуется интенсивным ростом очень тонких корней: биомасса и длина корней <0,5 мм на залежи по сравнению с сенокосом возросла в 2,1 и 1,7 раза соответственно (р < 0,001). Что, возможно, обусловлено изменением состава растительного сообщества, улучшением минерального питания за счет поступления опада.
Таким образом, наши исследования показали, что фракция тонких корней диаметром < 0,5 мм в верхнем слое почвы является наиболее чувствительной к изменению типов землепользования по сравнению с фракцией 0,5–2,0 мм, что подтверждается изменением морфологических признаков. На стадии самовосстановления нарушенных земель наблюдается интенсивное накопление биомассы корней <0,5 мм, а также увеличение их длины. В слое почвы 10–20 см тип землепользования не оказал существенного влияния на биомассу и длину корней всех изученных классов диаметра.
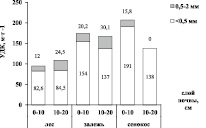
Рис. 3. Удельная длина корней по классам диаметра (<0,5 мм и 0,5-2,0 мм) и глубине почвенного слоя (0–10 см и 10–20 см)
Совместное изучение биомассы и длины корней отражает влияние условий среды обитания на развитие корней и, следовательно, на рост и развитие растительного сообщества. Удельная длина корней (УДК) является важнейшим параметром корневых систем, используемых в современной функциональной морфологии. Она показывает длину единицы сухой массы корня, оценивает затраты органических веществ, необходимых растению для образования единицы корней. В исследованиях Ф.С. Салпагаровой [9] значения удельной длины корня положительно коррелировали с содержанием азота и отрицательно с содержанием углерода в тонких корнях. Таким образом, растения с высокой удельной длиной корня образуют более «дешевые» корни, тратя на их построение меньше углерода. Увеличение УДК показывает «удешевление стоимости» корней, невысокое вложение продуктов фотосинтеза в их построение.
Исследование корней <0,5 мм на уровне растительных сообществ показало, что показатель УДК в естественной лесной экосистеме зоны южной тайги в слое почвы 0–10 см составил 83 м/г (рис. 3).
Вовлечение земель в сельскохозяйственный оборот увеличило УДК в 2,3 раза (p < 0,001). На стадии залежи УДК снизилась до 154 м/г, однако значимость различий не подтверждена статистически (p = 0,858). Полученные нами данные согласуются с результатами исследований I. Prieto и соавторов [10]. Они показали, что тонкие корни в растительных сообществах на нарушенных землях трех изучаемых климатических зон (тропической, средиземноморской, горной) обладали более высокой удельной длиной корней и противоположно вели себя в менее нарушенных экологических сообществах. УДК считается показателем интенсивности использования тонкими корнями одинакового объема почвы за счёт единицы потраченной биомассы [11]. В наших исследованиях УДК возрастает в ряду лес – залежь – сенокос, что свидетельствует о более интенсивном использовании почвы растениями сенокоса. Это может быть обусловлено как снижением плодородия в условиях активного землепользования, так и составом растительного сообщества, которое состоит из травянистых видов. Мы не обнаружили значительных изменений УДК корней 0,5–2,0 мм, связанных с типом землепользования. Предположительно, корни такого диаметра менее чувствительны к изменениям условий среды.
Заключение
Таким образом, наши исследования показали, что наибольшим изменениям в зависимости от состояния экосистемы подвержены морфологические признаки очень тонких корней <0,5 мм в верхнем слое почвы. На стадии самовосстановления нарушенных земель наблюдается интенсивное накопление биомассы корней данной фракции, а также увеличение их длины, что создает благоприятные условия для роста запасов органического углерода в почве. Возрастание удельной длины корней в ряду лес – залежь – сенокос свидетельствует о более интенсивном использовании объема почвы растениями сенокоса, что может быть обусловлено как уровнем плодородия, так и составом изучаемых растительных сообществ. Тем не менее, эти вопросы, требуют дальнейшего изучения.
Исследованиями I. Prieto и соавторов [10] показано, что тонкие корни агроэкосистем залежных земель разлагаются быстрее, чем корни фоновых ненарушенных сообществ. В нашем исследовании наибольшая биомасса тонких корней выявлена на залежи, что свидетельствует о стадии активного накопления органического вещества в постагрогенной экосистеме. Поэтому в дальнейших исследованиях необходимо установить количество и качество органического вещества, поступающего в почву при разложении тонких корней в связи с изменением землепользования в зоне южной тайги.
Библиографическая ссылка
Кондратова А.В., Абрамова Е.Р. ОСОБЕННОСТИ ФОРМИРОВАНИЯ ТОНКИХ КОРНЕЙ НА РАЗЛИЧНЫХ СТАДИЯХ ВОССТАНОВЛЕНИЯ ПОСТАГРОГЕННЫХ ЭКОСИСТЕМ В ЗОНЕ ЮЖНОЙ ТАЙГИ // Успехи современного естествознания. 2018. № 9. С. 18-22;URL: https://natural-sciences.ru/en/article/view?id=36860 (дата обращения: 20.06.2026).