


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
RESPONSE REACTION OF BARLEY IN ONTOGENESIS FOR POLLUTION BY COPPER

Медь является одним из биологически значимых и незаменимых микроэлементов для растений [1], однако при повышенных концентрациях может вызывать токсическое действие [2–4]. Медь, входя в состав ферментов, регулирует фотосинтез и концентрацию ингибиторов роста, водный обмен и перераспределение углеводов [5]. Недостаток меди вызывает у растений задержку роста [6]. Избыток меди вызывает ответную реакцию антиоксидантной защиты. В результате индуцируемого медью оксидативного стресса происходит усиленное образование свободных радикалов и активных форм кислорода, вызывающих перекисное окисление липидов (ПОЛ), инактивацию энзимов и нарушение в структуре ДНК. В реализации защитного потенциала растений от избытка тяжелых металлов (ТМ) важная роль отводится системе антиоксидантной защиты (синтез низкомолекулярных соединений, индукция пероксидаз, свободного пролина и т.д.). При высоком содержании меди в почве отмечается снижение роста и продуктивности как диких [7–9], так и культурных растений [10–12].
Исследования влияния меди на рост и различные биохимические показатели растений проводятся, как правило, в водной или гидропонной культуре, в упрощенных по сравнению с естественной средой обитания для большинства высших растений модельных системах. В литературе нет единого мнения о механизмах действия меди на растения при выращивании на различных типах почв. Эффективность действия ТМ на сельскохозяйственные культуры зависит от степени окультуренности почв, возраста растений и т.д. [13]. Наибольшее значение для формирования урожая растений имеет фаза их интенсивного роста, когда происходит закладка продуктивных органов и основное накопление биомассы растений.
Цель работы: изучение ответных реакций растений ячменя в онтогенезе на фазе интенсивного роста при воздействии различных концентраций меди.
Материалы и методы исследования
В вегетационном опыте объектом исследований служил ячмень Hordeum vulgare L., сорта Зазерский 85. Использовали два типа почв – дерново-подзолистую супесчаную и чернозем выщелоченный тяжелосуглинистый. Агрохимические характеристики почв составили соответственно: pHKCl 5,47 и 5,53; гумус – 1,7 % и 4,8 %; обменный K2O – 64,7 и 134,3 мг/кг; подвижный P2O5 – 805 и 214 мг/кг; гидролитическая кислотность – 2,7 и 3,0 мг-экв/100 г; сумма обменных оснований – 7,6 и 31,7 мг-экв/100 г. В контроле валовое содержание меди в почвах составляло 3,8 и 9,1 мг/кг. Для посева растений использовали вегетационные сосуды диаметром 22 см, высотой 20 см, с дренажем. За десять дней до посева медь добавляли в почву в виде водных растворов соли Cu(NO3)2 х 3H2O в концентрациях, соответствующих её содержанию: в дерново-подзолистой супесчаной почве – 50; 100; 150; 200 мг/кг; в черноземе выщелоченном тяжелосуглинистом – 100; 150; 300; 400 мг/кг, что с учетом валового содержания давало превышение ориентировочно допустимой концентрации (ОДК) [14] от 1,6 до 6,2 раза. Фон азота в разных вариантах опыта после добавления нитрата меди выравнивали внесением NH4NO3 до 200 мг/кг почвы по азоту. Полив растений осуществляли дистиллированной водой через дренажную трубку по мере высыхания почвы с контрольным взвешиванием сосудов. Ячмень выращивали в течение 60 суток. Плотность посева – 13 растений на сосуд, повторность опыта пятикратная.
Через 20, 30, 45 и 60 суток от даты всходов проводили анализ ростовых и биохимических показателей растений. Этапы органогенеза определяли по Ф.М. Куперман [15]. Массу надземной (без учета веса колоса) и корневой частей растений оценивали гравиметрическим методом для воздушно-сухого состояния. Спектрофотометрическим методом на приборе UNICO-1200 (Санкт-Петербург, РФ) определяли интенсивность ПОЛ по содержанию малонового диальдегида (МДА) и накопление свободного пролина согласно методике [16], повторность трехкратная. Данные в таблице приведены в виде средних и их стандартных ошибок. Достоверность различий вариантов устанавливали на основе двустороннего t-критерия для средних.
Результаты исследования и их обсуждение
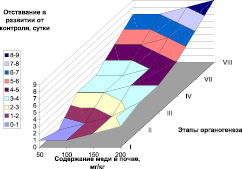
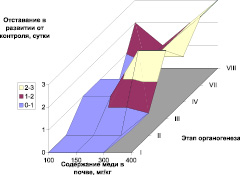
Следует отметить, что внесение меди в почву вызвало заметную задержку в развитии растений, по сравнению с контролем (рис. 1). Можно отметить прямую зависимость длительности органогенеза от содержания меди в почве, наиболее выраженную для дерново-подзолистой почвы. VIII этап органогенеза (колошение) в данном случае наступал на 7–9 дней позже, чем у контроля, и данное отставание в развитии накапливалось постепенно, во все этапы органогенеза. Наблюдаемые закономерности хорошо согласуются с динамикой накопления меди корнями и наземной биомассы данных растений [17]. Однако наблюдаемый эффект зависит не только от содержания меди в растениях, но и от типа почвы, возможно в данном случае проявление эффекта задержки развития обусловлено также количеством гумуса в почве или другими агрохимическими характеристиками почвы.
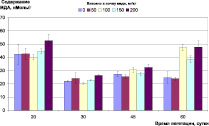
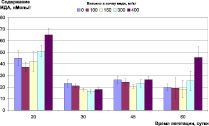
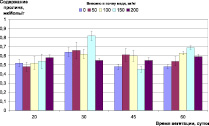
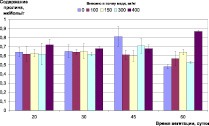
Контроль окислительно-восстановительного гомеостаза растений в ходе эксперимента показал, что внесение меди в почву в данных концентрациях не вызывает заметного стресса у ячменя (рис. 2). Содержание МДА в листьях на раннем этапе развития растений находилось в пределах, 40–60 нМоль/г, снизившись в дальнейшем до 20–30 нМоль/г, и не зависело от концентрации меди. На позднем этапе роста растений (фаза цветения – колошения) можно отметить достоверное (p < 0,05) увеличение (до 50 нМоль/г) содержания МДА в листьях растений, культивируемых на дерново-подзолистой почве при содержании меди в ней 100 мг/кг и выше, и на черноземе при концентрации меди 400 мг/кг (рис 2, А, Б).
А)
Б)
Рис. 1. Задержка в наступлении этапов органогенеза (по Ф.М. Куперман) в зависимости от валового содержания меди в дерново-подзолистой (А) почве и черноземе (Б)
А) Б)
В) Г)
Рис. 2. Окислительно-восстановительный гомеостаз растений ячменя на различных типах почв при внесении различных доз меди: А) содержание МДА, нМоль/г (дерново-подзолистая почва); Б) содержание МДА, нМоль/г (чернозем); В) содержание пролина, мкМоль/г (дерново-подзолистая почва); Г) содержание пролина, мкМоль/г (чернозем)
Концентрация пролина в листьях растений не зависела от содержания меди в почве и в течение всего эксперимента находилась в пределах 0,5–0,7 мкМоль/г (рис. 2, В, Г). Активизация в накоплении пролина у растений в ответ на неблагоприятные факторы является особенностью биосинтеза и метаболизма этого соединения для поддержания клеточного гомеостаза и может проходить независимо от возраста растений. Поэтому относительно стабильный и не зависимый от концентрации меди в растениях уровень пролина свидетельствует об отсутствии стресса растений и подтверждает, что закономерности накопления меди ячменем, представленные в работе [17], получены на фоне нормального физиологического состояния растений.
В таблице приведены данные динамики биомассы надземной и корневой части растений ячменя в онтогенезе. Для дерново-подзолистой почвы можно отметить рост надземной биомассы растений по сравнению с контролем при валовом содержании меди в почве в районе 50– 100 мг/кг и ее угнетение при уровне меди 200 мг/кг почвы. На черноземе эффекта увеличения надземной биомассы не наблюдалось, но отмечено достоверное ее снижение по сравнению с контролем на ранних (до 30 суток) и поздних (60 сутки) этапах роста растений, при этом выраженность эффекта зависела от содержания меди в почве и отмечалась при уровнях 300–400 мг/кг почвы.
Влияние меди на биомассу надземной и корневой части растений ячменя
|
Показатель |
Внесено меди в почву, мг/кг |
Время вегетации, сутки |
|||
|
20 |
30 |
45 |
60 |
||
|
Дерново-подзолистая супесчаная почва |
|||||
|
Надземная биомасса, г |
0 |
1,17 ± 0,09 |
1,76 ± 0,07 |
4,27 ± 0,04 |
9,96 ± 0,04 |
|
50 |
1,36 ± 0,02 |
2,19 ± 0,06* |
5,12 ± 0,04* |
11,18 ± 0,50 |
|
|
100 |
1,42 ± 0,11 |
2,16 ± 0,03* |
5,10 ± 0,05* |
14,4 ± 0,52* |
|
|
150 |
1,43 ± 0,01 |
2,02 ± 0,02* |
4,76 ± 0,07* |
9,62 ± 0,05 |
|
|
200 |
1,22 ± 0,06 |
1,57 ± 0,11 |
2,91 ± 0,03* |
7,94 ± 0,54* |
|
|
Масса корней, г |
0 |
0,18 ± 0,03 |
0,21 ± 0,02 |
0,43 ± 0,04 |
0,7 ± 0,06 |
|
50 |
0,18 ± 0,04 |
0,38 ± 0,05 |
0,39 ± 0,06 |
0,8 ± 0,04 |
|
|
100 |
0,18 ± 0,01 |
0,38 ± 0,01* |
0,44 ± 0,02 |
1,15 ± 0,09* |
|
|
150 |
0,24 ± 0,01 |
0,64 ± 0,07* |
1,16 ± 0,01* |
1,2 ± 0,01* |
|
|
200 |
0,26 ± 0,02 |
0,48 ± 0,11* |
1,36 ± 0,10* |
1,54 ± 0,20* |
|
|
Чернозем выщелоченный тяжелосуглинистый |
|||||
|
Надземная биомасса, г |
0 |
1,69 ± 0,02 |
2,68 ± 0,05 |
5,14 ± 0,18 |
13,5 ± 0,23 |
|
100 |
1,46 ± 0,05 |
2,46 ± 0,04 |
4,55 ± 0,08 |
14,3 ± 0,11 |
|
|
150 |
1,36 ± 0,10* |
2,36 ± 0,02* |
5,10 ± 0,28 |
10,6 ± 0,62 |
|
|
300 |
1,34 ± 0,02* |
2,15 ± 0,04* |
5,23 ± 0,16 |
9,03 ± 0,09* |
|
|
400 |
1,38 ± 0,07 |
2,14 ± 0,25* |
5,04 ± 0,07 |
8,56 ± 0,31* |
|
|
Масса корней, г |
0 |
0,18 ± 0,02 |
0,34 ± 0,01 |
0,53 ± 0,06 |
1,16 ± 0,02 |
|
100 |
0,17 ± 0,01 |
0,28 ± 0,01 |
0,44 ± 0,02 |
1,25 ± 0,11 |
|
|
150 |
0,16 ± 0,01 |
0,3 ± 0,02 |
0,48 ± 0,03 |
0,95 ± 0,08* |
|
|
300 |
0,16 ± 0,01 |
0,24 ± 0,02 |
0,42 ± 0,03 |
0,62 ± 0,08* |
|
|
400 |
0,19 ± 0,02 |
0,32 ± 0,01 |
0,55 ± 0,07 |
0,64 ± 0,03* |
|
Примечание. Различия с контролем значимы при: *p < 0,05.
Аналогичная картина снижения биомассы при уровнях содержания меди 150–400 мг/кг чернозема наблюдалась и для корневой системы ячменя на позднем этапе роста (60 сутки). В то же время на дерново-подзолистой почве с увеличением концентрации меди до 150–200 мг/кг происходит увеличение коневой биомассы, до 2–3 раз, по сравнению с контролем. Данный, зависимый от содержания меди в почве, рост корней растений ячменя возможно следует рассматривать как адаптивную реакцию, направленную на снижение концентрации меди в корневой системе за счет эффекта «разбавления». Однако на черноземе при тех же концентрациях меди в корнях ячменя [17] данный эффект не наблюдается.
Отмеченные возрастные особенности формирования надземной и корневой части биомассы ячменя в зависимости от концентрации меди на изученных типах почв, по-видимому, обусловлены различиями в барьерных функциях, развивающихся в условиях загрязнения ТМ почв, обладающих различными буферными свойствами и уровнем плодородия [13].
Выводы
1. Ответная реакция растений ячменя в ходе онтогенеза на возрастающие концентрации меди в почвах зависит от фазы развития и концентрации металла и в наибольшей степени проявляется на поздних этапах роста (фаза цветения и колошения).
2. Внесение меди в почву вызвало заметную задержку в развитии растений, по сравнению с контролем. Отмечена прямая зависимость длительности органогенеза от содержания меди в почве, наиболее выраженная для дерново-подзолистой почвы.
3. Контроль окислительно-восстановительного гомеостаза растений в ходе эксперимента показал, что внесение меди в дерново-подзолистую почву и чернозем в концентрациях соответственно до 200 и 400 мг/кг, не вызвал заметного стресса у ячменя. На позднем этапе роста растений (фаза цветения-колошения) можно отметить достоверное (p < 0,05) увеличение содержания МДА в листьях растений (с 20 до 50 нМоль/г), культивируемых на дерново-подзолистой почве при содержании меди в ней 100 мг/кг и выше и на черноземе при концентрации меди 400 мг/кг.
4. Особенности формирования надземной и корневой части биомассы ячменя в условиях повышенной концентрации меди в почве могут существенно зависеть не только от содержания меди в растениях, но и от типа почвы. Данные адаптивные реакции, по-видимому, обусловлены различиями в барьерных функциях, развивающихся в условиях загрязнения ТМ почв, обладающих различными буферными свойствами и уровнем плодородия.
Библиографическая ссылка
Цыгвинцев П.Н., Гончарова Л.И., Рачкова В.М. ОТВЕТНАЯ РЕАКЦИЯ ЯЧМЕНЯ В ОНТОГЕНЕЗЕ НА ЗАГРЯЗНЕНИЕ ПОЧВ МЕДЬЮ // Успехи современного естествознания. 2018. № 11-2. С. 305-310;URL: https://natural-sciences.ru/en/article/view?id=36944 (дата обращения: 13.07.2026).