


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
NFLUENCE OF ENVIRONMENTAL GENETIC FACTORS ON MORPHOMETRIC PARAMETERS AND DYNAMICS OF THE ANNUAL GROWTH OF WOOD IN PINUS SYLVESTRIS L. TEST CULTURES

Сведение и гибель лесов в результате рубок, пожаров и других видов воздействий, особенно за последние десятилетия, привели к очевидному сокращению их генетического разнообразия, значимых генотипов и оскуднению генофонда лесов Восточного Забайкалья. Сохранение оставшейся совокупности генных вариаций имеет серьезное значение для сбережения, рационального использования и репродукции экологически и экономически необходимых популяций, групп популяций и видов [1–3].
Все вышесказанное служит причиной продолжения ранее начатых работ по формированию постоянной лесосеменной базы сосны обыкновенной, которая является ценной лесообразующей породой данной территории. На ее долю приходится порядка 9 % лесопокрытой площади Восточного Забайкалья (по данным государственного лесного реестра на 01.01.2018 г.). Учитывая, что сосна является основным источником древесины исследуемой территории, этот вид может использоваться для создания плантаций на повышенную семенную и стволовую продуктивность. В связи с этим особую ценность представляют испытательные плантационные культуры, созданные путем размножения плюсовых деревьев (ПД), отобранных по ценным фенотипическим особенностям [4].
Существенным недостатком селекционной работы является то, что фенотипические признаки ПД не всегда соответствуют генотипу особи. Тем не менее отбор ПД по их фенотипу в какой-то мере оправдывается вероятностью того, что какая-либо ценная особенность этих деревьев проявится в их семенном потомстве значительно больше, чем в потомстве деревьев, не обладающих этой особенностью [5, 6].
Эффективность селекционных работ на плантациях, созданных путем вегетативного размножения (прививкой) ПД, снимает многие проблемы селекционно-генетической оценки, возникающие при испытании семенного потомства. Это связано с тем, что на ЛСП создаются сходные условия светового и минерального питания, вводятся повторности, клоны размещаются по определенной схеме. В результате здесь складываются условия, позволяющие фактически оценивать клоны ПД по целому ряду признаков [7–9], которые могут быть скрыты в высокополнотном естественном насаждении, на этапе отбора.
На территории Восточного Забайкалья первый опыт по отбору ПД и созданию лесосеменной плантации (ЛСП) путем их вегетативного размножения был проведен в начале 1990-х гг. сотрудниками лаборатории лесных экосистем Института природных ресурсов СО РАН (ныне ИПРЭК СО РАН) под руководством В.П. Бобринева и при непосредственном участии Л.Н. Пак. Ими было отобрано и аттестовано 52 ПД сосны обыкновенной в Читинском лесничестве. Здесь же была заложена первая опытная, клоновая плантация площадью 10 га.
Цель исследования: анализ клонового потомства ПД сосны обыкновенной по изменчивости морфометрических параметров и динамики годичного прироста древесины в испытательных культурах.
Материалы и методы исследования
Исследования проводились в 2014 г. на клоновой ЛСП сосны обыкновенной первого порядка (N 51 °49', Е 113 °08', 700 м над у.м.) в Читинском лесничестве на территории Восточного Забайкалья. Культуры создавались равномерной посадкой, с размещением клонов и повторностей (рамет) – 125 растений на 1 га плантации. Подробное описание закладки культур было приведено ранее [10, 11].
Морфометрические и дендрохронологические исследования выполнены согласно общепринятым методам [12, 13]. Изучению подлежали следующие морфометрические показатели: диаметр и высота ствола, возраст и длина хвои, плодоношение. Всего обследовано 21 клон ПД, 121 рамет. В качестве контроля были исследованы 14 непривитых, сопоставимых по возрасту деревьев сосны обыкновенной, произрастающих в сходных условиях за пределами ЛСП. Для определения динамики ширины годичных колец (ШГК) у каждого дерева по двум радиусам брали керны.
Результаты исследования и их обсуждение
Результаты статистических расчетов морфометрических данных приведены в таблице. К 30-летнему возрасту клоны имели среднюю высоту 7,4 м. По клонам данный показатель колебался от 6,8 до 8,1 м. Изменчивость высоты была небольшая, составляла всего 4,1 %. Наибольшая высота отмечалась у клонов: 24; 32; 37–38; 45, что охватывало только 23,8 % от числа всех клонов ПД. Клоны, не превышающие среднюю высоту по ЛСП, составляли 38 %.
Морфометрические параметры исследуемых деревьев
|
Параметр |
Контроль |
Кл_24 |
Кл_25 |
Кл_26 |
Кл_27 |
Кл_32 |
Кл_33 |
Кл_37 |
Кл_38 |
Кл_39 |
|
Диаметр ствола, см |
14,89 ± 1,19 (12,3–16,4) |
14,67 ± 1,56 (12,2–17,3) |
14,38 ± 2,06 (11,8–16,6) |
16,07 ± 2,35 (13,2–18,9) |
14,92 ± 3,81 (10,5–20,7) |
14,51 ± 2,1 (10,8–17) |
14,88 ± 2,5 (11,4–18) |
14,51 ± 2,1 (10,8–17) |
14,7 ± 1,52 (12,1–16,8) |
15,93 ± 1,96 (13,2–18,8) |
|
Высота ствола, м |
6,57 ± 0,44 (5,8–7,6) |
7,83 ± 0,39 (7,4–8,4) |
7,12 ± 0,62 (6,3–8) |
7,43 ± 0,77 (6,1–8,3) |
7,0 ± 1,09 (5,2–8,3) |
7,8 ± 0,84 (6,6–8,6) |
7,45 ± 0,55 (6,9–8,3) |
7,8 ± 0,84 (6,6–8,6) |
7,61 ± 0,78 (6,8–9) |
7,28 ± 1,01 (5,6–8,1) |
|
Плодоношение, шт. |
1,71 ± 0,83 (1–3) |
2,71 ± 0,76 (2–4) |
1,5 ± 1,05 (0–3) |
2.0 ± 0,89 (1–3) |
1,67 ± 0,82 (1–3) |
2,17 ± 1,17 (1–4) |
2,83 ± 0,98 (1–4) |
1,5 ± 0,84 (1–3) |
2,14 ± 1,07 (1–4) |
1,67 ± 1,21 (0–3) |
|
Возраст хвои, лет |
4,64 ± 0,74 (4–6) |
4,29 ± 0,49 (4–5) |
4,5 ± 0,84 (4–6) |
4,5 ± 0,55 (4–5) |
4,5 ± 0,55 (4–5) |
4,67 ± 0,52 (4–5) |
4,5 ± 0,84 (4–6) |
4,0 ± 0 (4) |
4,29 ± 0,76 (3–5) |
4,5 ± 0,55 (4–5) |
|
Длина хвои, см |
6,32 ± 0,98 (4,2–7,7) |
7,61 ± 1,23 (6–9,3) |
7,13 ± 0,67 (6,4–8,2) |
7,3 ± 1,31 (5,2–8,6) |
7,63 ± 1,4 (5,2–9,2) |
7,45 ± 0,86 (5,8–8,2) |
7,55 ± 0,86 (6,2–10,3) |
7,38 ± 1,24 (5,1–8,5) |
8,57 ± 0,98 (7,5–10,2) |
6,95 ± 1,02 (6–8,6) |
|
Выборка, шт. |
14 |
7 |
6 |
6 |
6 |
6 |
6 |
6 |
7 |
6 |
|
Окончание таблицы |
||||||||||
|
Диаметр ствола, см |
15,02 ± 0,93 (12,4–17,6) |
15,2 ± 0,84 (14,2–16) |
17,4 ± 3,77 (14,2–22) |
16,73 ± 3,07 (14,6–22) |
14,38 ± 1,66 (12,3–17,0) |
15,32 ± 2,3 (12–18) |
14,0 ± 1,2 (12,3–15,4) |
15,52 ± 2,15 (13,6–18,4) |
15,19 ± 1,55 (12,9–17) |
13,74 ± 1,02 (12,2–15) |
|
Высота ствола, м |
7,14 ± 0,91 (6,3–8,3) |
7,33 ± 0,85 (6,4–8,5) |
7,44 ± 0,76 (6,1–7,9) |
7,47 ± 0,64 (6,6–8,1) |
7,27 ± 0,56 (6,7–7,9) |
8,1 ± 0,24 (7,8–8,5) |
6,82 ± 1,04 (5,1–7,6) |
7,55 ± 0,77 (6,3–8,3) |
7,27 ± 0,56 (6,5–8,3) |
7,47 ± 0,74 (6,4–8,6) |
|
Плодоношение, шт. |
2,2 ± 0,84 (1–3) |
1,67 ± 0,82 (1–3) |
3 ± 1,22 (1–4) |
2,17 ± 1,17 (1–4) |
2,83 ± 0,41 (2–3) |
2,33 ± 1,20 (1–4) |
1,2 ± 0,45 (1–2) |
2,5 ± 0,55 (2–3) |
1,57 ± 0,53 (1–2) |
2,14 ± 0,9 (1–3) |
|
Возраст хвои, лет |
4,2 ± 0,45 (4–5) |
4,5 ± 0,55 (4–5) |
4,0 ± 0,71 (3–5) |
4,67 ± 0,82 (4–6) |
4,67 ± 0,52 (4–5) |
4,0 ± 0,63 (3–5) |
4,4 ± 0,55 (4–5) |
4,5 ± 0,84 (4–6) |
4,71 ± 0,49 (4–5) |
4,29 ± 0,49 (4–5) |
|
Длина хвои, см |
6,88 ± 1,46 (4,5–8,5) |
8,23 ± 0,6 (7,4–8,9) |
7,58 ± 1,64 (4,7–8,7) |
7,03 ± 0,69 (6,1–7,8) |
7,75 ± 0,87 (6,9–9,1) |
7,78 ± 0,95 (6,5–9,1) |
7,56 ± 0,94 (6,1–8,7) |
7,87 ± 1,28 (6,4–9,9) |
6,74 ± 1,49 (4,8–8,6) |
8,24 ± 0,79 (7,4–9,3) |
|
Выборка, шт. |
5 |
6 |
5 |
6 |
6 |
6 |
5 |
6 |
7 |
7 |
Примечание. В скобках приведены минимальные и максимальные значения параметров.
Анализ морфометрических данных клонов ПД и контрольных деревьев показал, что средняя высота клонов была достоверно больше (от 6,82 ± 1,04 м до 8,1 ± 0,24 м), чем у непривитых деревьев (6,57 ± 0,44 м).
Анализ динамики ширины годичных колец (ШГК). По результатам измерения средний диаметр клонов в культурах составил 15,1 см. Изменчивость этого показателя была небольшой – 6,1 %. По диаметру, наибольшее превышение среднего показателя по культурам, отмечалось у клонов: 26; 42–43. Клоны, не превышающие средний диаметр по ЛСП, составляли 47,6 %. Средний диаметр клонов был достоверно больше (от 13,74 ± 1,02 см до 17,4 ± 3,77 см), чем у непривитых деревьев (14,89 ± 1,19 см).
Длина хвои у клонов изменялась незначительно, уровень изменчивости составил 6,5 %. Длинную хвою имели клоны: 38; 41; 52, т.е. 14,2 % всех клонов. Клоны, имеющие хвою меньше средней по ЛСП, составляли 38 %. У контрольных деревьев в среднем длина хвои достоверно отличалась от клонов, она составляла 6,32 ± 0,98 см, в то время как у клонов варьировала от 6,74 ± 1,49 см до 8,57 ± 0,98 см.
Небольшая часть деревьев из исследуемой выборки характеризовалась лучшим плодоношением (14,2 %). Это клоны 24; 42; 44. Изменчивость данного показателя была существенной 24,7 %. Лишь у 38,0 % деревьев данный показатель был сопоставим с контролем.
Возраст хвои не имел значимых различий по сравниваемым выборкам, это обусловлено тем, что данный параметр не зависит от генетической предрасположенности, а связан с влиянием экологических факторов среды.
По результатам измерения ширины годичных колец построены 28 индивидуальных древесно-кольцевых хронологий, их продолжительность составила от 12 до 15 лет (рис. 1). Они показали, что для всех исследуемых приростов характерно формирование наиболее широких годичных колец в 2002, 2005, 2008 и 2009 гг., а узких – в 2001, 2003, 2007 и 2011 гг. Что согласуется с данными по исследуемой территории, полученными ранее [14, 15]. Для всех кернов характерно наличие минимальных экстремумов прироста в 2003 и 2007 гг. Сравнительный анализ средних значений ШГК у клонов ПД не показал достоверных отличий размеров годичных колец по сравнению со средней ШГК контрольных деревьев.
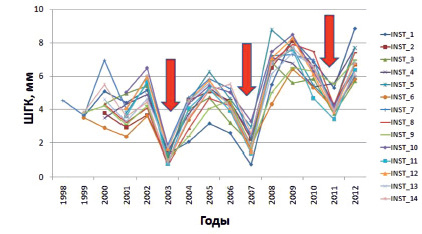
Рис. 1. Индивидуальные серии ширины годичных колец на клоновой ЛСП сосны обыкновенной первого порядка (среднее по двум радиусам)
За период исследования по всем сериям средние значения ШГК варьировали в пределах от 3,8 ± 1,64 мм до 5,4 ± 1,83 мм, максимальный прирост составил 8,9 мм, а минимальный – 0,8 мм. Такие высокие значения ШГК связаны, прежде всего, с возрастом деревьев, для которых, так называемый, «период большого роста», еще не завершен. Дисперсия по выборкам колебалась от 2,47 до 5,97.
Статистический анализ в программе TSAPWin показал высокое сходство индивидуальных графиков (Glk) и максимальный уровень синхронности (GSL) между ними. Среднее значение межсериального коэффициента корреляции составило – 0,97, при минимальном – 0,87, что позволило усреднить данные и получить обобщенную древесно-кольцевую хронологию INST по абсолютным значениям ШГК, отражающую влияние внешних факторов среды на радиальный прирост деревьев.
Анализ климатического отклика выполнен с помощью корреляционного анализа между полученной обобщенной древесно-кольцевой хронологией и среднемесячными значениями атмосферных осадков за период, в течение которого возможно их влияние на радиальный прирост древесины. Использованы данные метеорологической станции г. Чита (N 52 °03', Е 113 °29'). Доверительный интервал коэффициентов корреляции (при р < 0,05) лежит за пределами значений от 0,46 до –0,46.
Расчет показал, что значимое влияние на радиальный прирост деревьев, произрастающих на клоновой ЛСП сосны обыкновенной первого порядка, оказывают среднемесячные атмосферные осадки мая и июня (R = 0,62 и R = 0,61 соответственно) (рис. 2). Одинаковые значения коэффициента корреляции (R = 0,61) получены для годовой суммы осадков и суммы осадков за период вегетации (с мая по сентябрь), что связано с выпадением подавляющего количества атмосферных осадков в исследуемом районе в теплый период года.
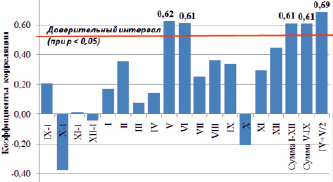
Рис. 2. Коэффициенты корреляции между обобщенной древесно-кольцевой хронологией (INST) и атмосферными осадками за период с 1999 по 2012 г.
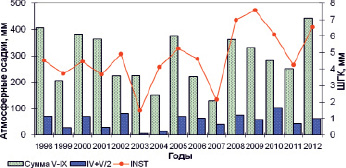
Рис. 3. Динамика приростов обобщенной древесно-кольцевой хронологии и значений атмосферных осадков за отдельные периоды
Несмотря на сходные показатели значимых коэффициентов корреляции для исследуемых периодов, при сравнении динамики размеров ШГК и атмосферных осадков было отмечено, что для отдельных лет определяющее значение оказывают, прежде всего, осадки первой половины периода вегетации (рис. 3). Например, в 2002 г., выявлены высокие значения прироста (4,9 мм), несмотря на то, что сумма осадков периода с мая по сентябрь была на уровне 2003 г. (223 и 224 мм), когда прирост был экстремально низким (1,5 мм). Это связано с тем, что в 2002 г. атмосферные выпадения в первой половине вегетации были почти в 16 раз выше по сравнению с тем же периодом в 2003 г. Подобная ситуация отмечается и для 2010 г. Поэтому в корреляционный анализ дополнительно были включены средние значения суммы атмосферных осадков июля и июня, которые позволили выявить максимальные значения показателя (R = 0,69). Достоверной связи приростов с атмосферными осадками сентября предшествующего года не выявлено (R = 0,21).
Заключение
Из полученных предварительных результатов следует, что у 57,8 % клонового потомства ПД сохраняется генетическая обусловленность по скорости роста. Несмотря на то, что по некоторым показателям не у всех клонов ПД выявлены достоверные отличия с контрольными деревьями, достоверно большая площадь ассимиляционной поверхности привитых деревьев позволяет заключить, что они характеризуются большей энергией протекания физиологических процессов, которые определяют формирование шишек и темпы образования биомассы. Погодичная динамика ШГК характеризуется наличием минимальных экстремумов прироста в 2003 и 2007 гг., связанных со снижением количества атмосферных осадков в эти годы. Разницы между деревьями по ШГК выявлено не было, предположительно это связано с возрастом деревьев и периодом большого роста. Оценка клонов ПД, произрастающих на ЛСП сосны обыкновенной первого порядка, с помощью дендрохронологических методов исследования показала, что древостои II класса возраста испытывают интенсивное воздействие природных факторов среды, которое выражается в формировании экстремально низких значений ширины годичных колец в годы засух. Особенно губительно на деятельности камбия сказывается снижение атмосферных осадков в первой половине периода вегетации (май – июнь).
Библиографическая ссылка
Вахнина И.Л., Ларин В.С., Пак Л.Н. ВЛИЯНИЕ ЭКОЛОГО-ГЕНЕТИЧЕСКИХ ФАКТОРОВ НА МОРФОМЕТРИЧЕСКИЕ ПАРАМЕТРЫ И ДИНАМИКУ ГОДИЧНОГО ПРИРОСТА ДРЕВЕСИНЫ В ИСПЫТАТЕЛЬНЫХ КУЛЬТУРАХ PINUS SYLVESTRIS L. // Успехи современного естествознания. 2020. № 1. С. 5-11;URL: https://natural-sciences.ru/en/article/view?id=37314 (дата обращения: 07.07.2026).
DOI: https://doi.org/10.17513/use.37314