


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
ECOLOGICAL AND GEOGRAPHICAL FEATURES OF FOREST FORMATION FORMATION IN THE MIDDLE CURRENT OF THE LOWER TUNGUSKA IN LAST LATE TIME

Голоцен (последние 10 тысяч лет от современности) является одной из более-менее разработанных моделей для расшифровки закономерностей естественного развития лесных экосистем прошлого. Однако определение влияния различных факторов на их формирование требует еще детального, на видовом уровне, изучения. Имеющийся к настоящему времени палинологический материал по голоцену Восточной Сибири малочисленный, интерпретация которого выполнена в основном на родовой таксономии ископаемых флор [1; 2]. Поэтому оценка особенностей влияния эколого-географических условий на организацию фитоценотической структуры регионального растительного покрова, как одного из компонентов геосистем, является определенным информационным вкладом в повышение качества реконструкции и прогноза изменений природной среды. Детальное воссоздание видовой структуры лесных сообществ, их эколого-ценотическая оценка позволят установить характер вековых сукцессионных процессов, скорость и направленность, величину сдвига природных зональных и провинциальных границ и миграционные возможности основных лесообразующих пород. Такая информация выявляет не только динамику пространственной дифференциации типов растительности, но и механизмы трансформации их структуры под воздействием различных внешних факторов в долговременной ретроспективе, и в первую очередь климата. И здесь применение палеокарпологического метода при изучении голоценовых отложений приобретает неотъемлемую часть палеоботанических исследований. Данные палеокарпологии дают таксономически наиболее богатые видовые палеокомплексы растений [1], которые лучше отражают условия среды.
Основная цель исследования – выполнить реконструкцию растительного покрова на основе палеокарпологического изучения голоценовых отложений. В ее задачи входило: выявить последовательность смен эколого-ценотических групп прошлых растительных сообществ, дать количественную оценку климата их существования и провести анализ динамики лесных формаций по гипсометрическим уровням.
Материалы и методы исследования
В работе представлены результаты палеокарпологического исследования двух разрезов, заложенных на двух тестовых участках: западный – разрез «Порох», восточный – разрез «Бугарикта». Их территории расположены в среднем течении р. Нижней Тунгуски средней подзоны тайги в пограничной полосе с северной тайгой.
Материалом для исследования стали макроостатки ископаемых растений (семена, плоды, вегетативные части растений, обломки и угольки древесины и др.), извлеченные из голоценовых отложений. Данные видового состава макроостатков доминантных видов растений в каждом разрезе позволили разделить их на макрокомплексы по лидирующим эдификаторам. Выделенные макрокомплексы – это временные интервалы, каждый из которых характеризуется своим ландшафтно-климатическим режимом.
Процедура обработки палеокарпологических данных проводилась с позиций лесоведения по общепринятой методике, с дополнениями авторов применительно к голоцену, что подробно освещалось авторами ранее [3]. Уместно отметить, что объектами палеокарпологии становятся только те остатки видов растений, которые были типичны и доминировали в прошлых фитоценозах. Особенностью применяемого авторами метода является то, что индивидуально изучается анатомо-морфологический портрет каждого ископаемого объекта. Поскольку голоценовым макроостаткам свойственна хорошая физиономичность, то помимо видовой идентификации это позволило дифференцировать их на три группы удаленности от материнского растения: локальная, сопредельная, региональная. Это хорошо диагностируется по наличию следов транспортировки на поверхности ископаемых объектов. По этому же признаку установлено, что во всех макрокомплексах остатки вегетативных частей растений по большей части являются локальными. На этой основе определялось пространственное распределение палеосообществ разной структуры и статуса. Судя по составу изученных ранее субрецентных макрокомплексов в центральной части Эвенкии (район п. Тура) [3], основная часть доминирующих и содоминирующих остатков каждого макрокомплекса в первую очередь правильно отражает растительное сообщество в точке его формирования. Содержание в комплексах остатков от 1 до 10 % указывает на состав растительности, окружающей точку исследования, характеризуя лесную формацию. Участие же в комплексе остатков менее 5 % и единично говорит об их привнесенности и свидетельствует о доминантах палеоландшафта. Таким образом, каждый ископаемый макрокомплекс позволяет реконструировать не только локальный тип растительности, но и местный, и региональный. Выявление эдификаторов и доминантов в прошлых растительных сообществах разного уровня проводилось с использованием метода эколого-ценотического анализа [4], применяемого при фитоиндикации современных лесных экосистем. И здесь, в первую очередь, наиболее четким показателем преобразований, происходящих в растительном покрове, являлся видовой состав травяно-кустарничкового яруса, как наиболее мобильного структурного компонента, быстрее реагирующего на различные флуктуации природных условий. По результатам анализа ископаемого материала для каждого разреза построены карпограммы, отражающие изменения видового состава комплексов макроостатков растений по временным отрезкам голоцена, а, следовательно, и динамику биоразнообразия прошлых фитоценозов (например, рис. 1).
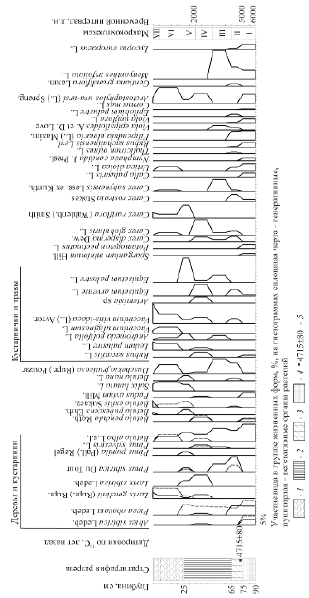
Рис. 1. Карпограмма разреза «Порох» Виды торфа: 1 – осоково-сфагновый, 2 – древесный, 3 – гипново-древесный. 4 – Песок. 5 – Глубина отбора образцов на радиоуглеродный анализ и датировки по 14С
Для характеристики палеоклимата в точках исследования привлечены данные метеостанций аналоговых территорий. Основным актуалистическим источником палеореконструкций стали геоботанические карты современного растительного покрова, лесоустроительные материалы, а также при заключительном анализе палеоботанического материала стала эколого-фитоценотическая карта современного растительного покрова Сибири.
Результаты исследования и их обсуждение
Западный тестовый участок расположен на правом берегу р. Нижняя Тунгуска надпойменной террасы приустьевой части р. Порох в междуречье рек Кочумдек и Тутончана. В растительном покрове здесь доминируют лиственничники зеленомошно-лишайниковые и кустарничково-моховые с примесью березы, ели, кедра и реже сосны. Закладка разреза «Порох» (64 °24'27" с.ш., 93 °4'48" в.д.) и отбор образцов сделаны в редкостойном лиственничнике (9Л1Б) осоково-багульниково-сфагновом в окружении лиственничника лишайниково-зеленомошного с небольшой примесью березы и кедра. На основании палеокарпологического анализа образцов построена карпограмма (рис. 1) эколого-ценотических спектров комплексов макроостатков растений по временным отрезкам голоцена, которые отразили динамику трансформации растительного покрова начиная с 6000 лет назад (далее л.н.). В первой половине периода на территории господствовали пихтово-еловые и елово-кедровые, в конце с участием сосны, разнотравные леса южнотаежного облика. Далее их сменили лиственничные формации, вначале с доминированием лиственницы сибирской, затем с лиственницей Гмелина.
Восточный тестовый участок расположен на левом берегу Нижней Тунгуски в 3,5 км на ССЗ от устья р. Бугарикта (200 км выше п. Тура). На территории наиболее высокие части водоразделов заняты лиственнично-сосновыми бруснично-багульниково-моховыми лесами с подлеском из ольхи. На менее высоких и пониженных частях господствуют разные типы кустарничково-сфагновых лиственничников. Большие площади заняты верховыми кустарничково-сфагновыми болотами. Разрез «Бугарикта» (64 °0'46" с.ш., 103 °4'27" в.д.) заложен на осоково-багульниково-сфагновом болоте в окружении лиственничника багульниково-брусничного с елью, который выше по уровню сменяется лиственнично-сосновым лесом толокнянково-бруснично-лишайниковым. Палеокарпологический материал, представленный в карпограмме (рис. 2), отразил смены растительности, начиная с 7000 л.н. Здесь четко проявилось господство елово-лиственничных формаций с лиственницей сибирской в первой половине периода и с лиственницей Гмелина – во второй.
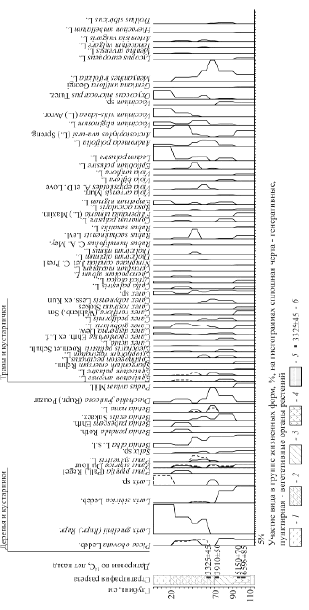
Рис. 2. Карпограмма разреза «Бугарикта», высота над у.м. – 170 м Виды отложений. Торф: 1 − сфагновый, 2 – сфагново-осоковый, 3 – осоковый, 4 – травяно-древесный. 5 – Песок. 6 – Радиоуглеродные датировки по14С
Общий таксономический состав реконструированной ископаемой флоры составил 70 таксон, из них – 65 виды.
На основании синтеза палеокарпологического материала, рассмотренного выше нового и полученного ранее по анализу разрезов лесных почв в окрестностях п. Туры [3], считается, что эволюция растительного покрова в среднем течении р. Нижней Тунгуски восстанавливается по семи возрастным уровням (таблица), начиная со второй половины атлантического периода общепринятой хроностратиграфической схемы голоцена [5].
Динамика голоценовых растительных формаций в бассейне р. Нижней Тунгуски в голоцене
|
Временные периоды голоцена, лет назад |
Время, датированное по 14С, лет назад |
Абсолютная высота над уровнем моря в метрах |
|
|
100 (западная часть) |
200 (восточная часть) |
||
|
Современность |
Лиственничная лишайниково-зеленомошная, ед. береза, кедр |
Лиственнично-сосновая толокнянково-бруснично-лишайниковая |
|
|
0–500 |
510 ± 30* |
Березово-лиственничная (Larix gmelinii) толокнянково-брусничная |
Березово-лиственничная (Larix gmelinii) кустарничково-моховая |
|
0–1000 |
910 ± 40# |
Лиственнично-кедровая (Larix gmelinii) бруснично-толокнянковая |
Кедрово-лиственничная (Larix gmelinii) толокнянково-брусничная с кедровым стлаником (Pinus pumila) |
|
1000–2000 |
1690 ± 40* |
Сосново-кедровые толокнянково-брусничная (Larix gmelinii, Larix sibirica) с кедровым стлаником (Pinus pumila) Т °янв выше на 3 °С, Т °июля выше на 2 °С. Годовые осадки больше на 60 мм |
Сосново-лиственничная кустарничковая (Larix gmelinii, Larix sibirica) |
|
2000–3000 |
2420 ± 45* |
Сосново-лиственничная бруснично-толокнянковая (Larix gmelinii, Larix sibirica) Т °янв ниже на 2 °С, Т °июля ниже на 1 °С. Годовые осадки меньше на 100 мм |
Сосново-лиственничная мохово- кустарничковая (Larix gmelinii, Larix sibirica) Т °янв ниже на 4 °С, Т °июля ниже на 0,5 °С. Годовые осадки меньше на 30 мм |
|
3000–4000 |
3325 ± 45* 3910 ± 80* |
Сосново-кедровая кустарничково-осоковая |
Березово-кедрово-лиственничная с ольхой кустарничковая (Larix sibirica) |
|
4000–5000 |
4715 ± 80# |
Елово-кедровая разнотравно-брусничная |
Елово-лиственничная (Larix sibirica) разнотравно-кустарничковая леса |
|
5000–6500 |
5150 ± 70# 6595 ± 85* |
Пихтово-еловая кустарниково-разнотравная южнотаежного облика Т °янв выше на 4 °С, Т °июля выше на 2 °С. Годовые осадки меньше на 50 мм |
Кедрово-лиственничная (Larix sibirica) разнотравно-кустарничковая с южнотаежными элементами леса в сочетании с лесостепными участками Т °янв выше на 2 °С, Т °июл выше на 1 °С. Годовые осадки больше на 150 мм |
Примечание. Радиоуглеродные датировки выполнены: * – в Институте леса им. В.Н. Сукачева СО РАН [1], # – в Институте геологии и минералогии им В.С. Соболева СО РАН к.г.-м.н. Л.А. Орловой.
Проведенные исследования выявили существование в конце атлантического периода мягкого, более сухого и теплого климата, чем ныне. В то время территория среднего течения Нижней Тунгуски была занята в западной части темнохвойной тайгой южнотаежного облика, а в восточной господствовали смешанные леса с участием южнотаежных элементов. Далее сформировавшаяся климатическая обстановка вследствие наступившего короткопериодного похолодания способствовала большему развитию смешанных среднетаежных лесов, исчезновению пихты, но появлению сосны и укреплению позиции кедра на западе. Позже пришедшее похолодание и нарастание континентальности климата привело к вытеснению темнохвойных пород и господству лиственничных формаций вначале с доминированием Larix sibirica, а затем только из Larix gmelinii. В последнем 2000-летнем интервале (после позднеголоценового похолодания 3000– 2000 л.н.) наступившее потепление и уменьшение континентальности климата способствовало сокращению участия лиственниц в древесном пологе лесных формаций, особенно Larix gmelinii в западной части бассейна р. Н. Тунгуска, и укреплению позиций сосны и кедра. В это же время в подлеске принимает участие и Pinus pumila, ареал которого постепенно смещается на восток. Последующее похолодание и усиление континентальности постепенно преобразовало растительный покров в современный облик.
Заключение
Установлено, что в динамике лесных формаций среднего течения р. Нижней Тунгуски в голоцене проявились общие закономерности эволюции растительного покрова Центральной части Средней Сибири, выявленные по палинологическим данным. Кроме того, корреляция полученного палеокарпологического материала с подобным палеоботаническим показывает четкое совпадение основных рубежей смен растительного покрова с хорошо датированными записями изменения природной среды Сибири [6; 7]. Выполненные исследования пополнили банк данных по видовому биоразнообразию растительности голоцена детально-региональной информацией мало изученной истории голоцена Эвенкии, что поможет надежнее строить научный прогноз для решения вопросов рационального природопользования.
Работа выполнена при поддержке РФФИ – грант № 18-04-01068.
Библиографическая ссылка
Кошкаров А.Д., Кошкарова В.Л. ЭКОЛОГО-ГЕОГРАФИЧЕСКИЕ ОСОБЕННОСТИ ФОРМИРОВАНИЯ ЛЕСНЫХ ФОРМАЦИЙ В СРЕДНЕМ ТЕЧЕНИИ НИЖНЕЙ ТУНГУСКИ В ПОСЛЕЛЕДНИКОВОЕ ВРЕМЯ // Успехи современного естествознания. 2021. № 2. С. 100-106;URL: https://natural-sciences.ru/en/article/view?id=37581 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/use.37581