


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
WATER REGIME OF SEA BUCKTHORN (Hippophae rhamnoides) OF THE ISSYK-KUL LAKE COAST

Древесно-кустарниковые растения, растущие в естественных условиях, выполняют огромную средообразующую, водоохранную и множество других функций. Произрастая в различных условиях, они адаптируются к условиям окружающей среды, вырабатывая своеобразные адаптивные механизмы, позволяющие им благополучно существовать. Но изменения, происходящие в природе, зачастую не носят скоростного характера, и поэтому растения успевают выработать адаптивные механизмы [1]. Изменения же, провоцируемые человеком, часто имеют динамичный характер, тем самым интенсивно воздействуя на экосистемы прямым и косвенным путем. Влиянию техногенных факторов на рост и развитие древесно-кустарниковой растительности, произрастающих в урбосистемах, природных парках, лесах, посвящено довольно много исследований [2]. В том числе много работ посвящено изучению морфологического, биохимического, генетического видоизменения различных частей растений при разных типах антропогенного воздействия [3, 4]. Произрастая и адаптируясь к различным неблагоприятным факторам окружающей среды, они успешно могут быть использованы для оценки качества окружающей среды. Одним из информативных параметров условий техногенного загрязнения окружающей среды является изменение водного режима и ассимиляционной активности растений [5, 6]. Как известно, водный режим древесно-кустарниковой растительности зависит от внутренних параметров растений и от факторов окружающей среды [7]. Исследователями установлено, что устойчивость растений к неблагоприятным факторам окружающей среды зависит от содержания и состояния воды в клетках и тканях растений [8, 9]. Это позволяет дифференцировать их как устойчивые и неустойчивые виды к техногенным изменениям окружающей среды. У устойчивых видов растений различные загрязнения природной среды приводят к увеличению количества связанной и свободной воды и к повышению их водоудерживающей способности. Для оценки степени антропогенного воздействия на прибрежные экосистемы большой интерес вызывают кустарниковые заросли, растущие на побережьях рек, озер и различных водоемов. Они выполняют огромную природоохранную роль, в том числе выполняя роль механического фильтра воды, укрепляя берега и почвы. Зачастую они подвергаются большому антропогенному прессингу: вырубке, перевыпасу скота, ареалы их распространения вычищаются для хозяйственно-бытового использования. С каждым годом растет антропогенное воздействие на уникальную экосистему озера. Растет количество рекреационных учреждений как отвечающих требованиям санитарно-эпидемиологического надзора, так и не соответствующих им. В настоящее время экологическое состояние облепиховых зарослей на побережье озера Иссык-Куля неудовлетворительное. Как известно, самоочищающаяся функция любого природного объекта имеет свой предел, и не всегда известно, когда этот предел будет перейден. С связи в этим экологическое состояние воды и экосистем, обрамляющих озеро Иссык-Куль, является одним из вопросов, вызывающих научный интерес ученых разных специальностей. Нами исследовались возможности оценки влияния антропогенного фактора на экосистему озера по состоянию водного режима кустарниковых зарослей облепихи крушиновидной, которая произрастает вокруг всей акватории озера, образуя местами непроходимые заросли. В связи с этим целью наших исследований являлось изучение водного режима облепиховых зарослей побережья озера Иссык-Куль при различном уровне антропогенного воздействия на экосистему озера – от повышенной до умеренной.
Материалы и методы исследования
Объектом наших исследований были дикорастущие заросли облепихи крушиновидной побережья озера Иссык-Куль. Отбор проб был произведен в пяти точках (1 – Оттук, 2 –Тамчы, 3 – Чон-Сары-Ой, 4 – Чолпон-Ата, 5 – Бостери) (рис. 1). Пункт сравнительно неосвоенного в рекреационном плане участка озера Оттук был взят в качестве контроля.
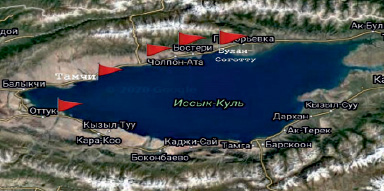
Рис. 1. Пункты отбора образцов
Пробные площади закладывались в ме- стах ближайшего расположения естественных зарослей облепихи крушиновидной к побережью воды озера Иссык-Куль. Лиcтья были отобраны из cpeднeй чacти хорошо освещенных пoбeгoв и побегов, которые pacпoлoжeны нa пepифepии cpeднeй чacти кpoны. Cвeжecoбpaнныe лиcтья зaвopaчивaли вo влaжную флaнeлeвую ткaнь и зaтeм пoмeщaли в пoлиэтилeнoвый пaкeт для выpaвнивaния вo вpeмeни экcпepимeнтa [10].
Для изучения водного режима растений определялись следующие показатели, представленные в табл. 1.
Таблица 1
Показатели водного режима растений
|
Определяемый параметр |
Характеристика |
Формула |
|
Интeнcивнocть тpaнcпиpaции |
Oпpeдeлена мeтoдoм быcтpыx взвeшивaний лиcтьeв, пepecчeтoм нa 1 г cыpoгo вeca зa 1 чac. Лиcтья взвeшивaлись двaжды: 1 – вес дo oпытa, 2 – вес чepeз 3 мин oбeзвoзживaния в кoмнaтныx уcлoвияx пpи тeмпepaтуpe 27 °C. Пo peзультaтaм взвeшивaния paccчитывaлacь пoтepя вoды в мг зa 1 чac нa 1 г cыpoй мaccы и нa eдиницу плoщaди (100 cм2) лиcтa. Пробы были отобраны в обеденное время, в августе 2020 г. Пoвтopнocть oпытa 6–8-кpaтнaя |
ИТ – интeнcивнocть тpaнcпиpaции, мг/г/чac; P1 – нaчaльный вec лиcтa, мг; P2 – вec лиcтa чepeз 3 мин, мг |
|
Содержание воды в листьях |
Лиcтья выcушивaли в тepмocтaтe дo пocтoяннoгo вeca пpи 105 °C |
CВ – coдepжaниe вoды, %; P0 – нaчaльный вec лиcтa, мг; P2 – aбcoлютнo cуxoй вec, мг |
|
Водопотеря |
Рacчeт вoдoпoтepи пpoвoдили в зависимости от пepвoнaчaльной cыpой мaccы лиcтa |
ВП – вoдoпoтepя, %; P0 – нaчaльный вec лиcтa, мг; P1 – вec чepeз 1 ч выcушивaния, мг |
|
Водный дефицит |
Сpeзaнныe лиcтья взвeшивали нa тopcиoнныx вecax и зaтeм пoгpужaли их в вoду кoмнaтной тeмпepaтуpы, нa 2 ч. Пocлe того, как листья нacыщались вoдoй, их cлeгкa пpocушивaли фильтpoвaльнoй бумaгoй и пoвтopнo взвeшивaли |
ВД – вoдный дeфицит, %; P0 – пepвoнaчaльный вec лиcтa, мг; P1 – вec лиcтa чepeз 2 ч, мг |
|
Засухоустойчивость |
Свeжecpeзaнныe лиcтья взвeшивaли нa тopcиoнныx вecax и oбeзвoживaли в тeчeниe 1 ч в кoмнaтныx уcлoвияx пpи тeмпepaтуpe 27 °C |
КЗУ – кoэффициeнт зacуxoуcтoйчивocти; P1 – мacca лиcтa дo выcушивaния, мг; P2 – мacca лиcтa пocлe выcушивaния, мг |
|
Жароустойчивость |
Определяли высушиванием листьев при температуре 40 °С в термостате в течение 1 ч, которые были предварительно взвешены на торсионных весах. После 1 ч экспозиции листья повторно взвешивались |
КЖУ – кoэффициeнт жapocтoйкocти; P1 – мacca лиcтa дo oпытa, мг; P2 – мacca лиcтa пocлe oпытa, мг |
|
Солеустойчивость |
Листья предварительно взвешивались на торсионных весах, далее помещались в чашки Петри с 10 % раствором хлористого натрия на 24 ч. После чего листья вновь взвешивались |
КCУ – кoэффициeнт coлeуcтoйчивocти; P1 – мacca лиcтa дo пoгpужeния в pacтвop, мг; P2 – мacca лиcтa пocлe пoгpужeния в pacтвop, мг |
Таблица 2
Интенсивность транспирации опытных образцов
|
№ п/п |
Пункты отбора |
Cpeднee знaчeниe, М ± m, мг/г/чac |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
6800 ± 14,83 |
4,38 |
|
2 |
Бостери |
6800 ± 14,83 |
4,38 |
|
3 |
Чолпон-Ата |
10600 ± 18,96 |
6,22 |
|
4 |
Тамчы |
6850 ± 12,52 |
4,11 |
|
5 |
Оттук |
6000 ± 13,45 |
4,24 |
Результаты исследования и их обсуждение
Интенсивность транспирации. Максимальная интенсивность транспирации отмечена в пункте Чолпон-Ата, составив 10 600 мг/г/час, а минимальный показатель был в пункте Оттук (табл. 2). Схожие показатели, равные 6850 ± , были в пунктах Булан-Соготту и Бостери. Интенсивность транспирации в наиболее часто посещаемом туристами пункте была наиболее высокой, а контрольный вариант Оттук имел наименьший показатель 6000 мг/г/час. Учитывая, что на побережье озера в летнее время стихийно распологаются летние кафе, вследствие чего вечерний пляж наполняется дымом и гарью, а также кустарники облепихи зачастую используются в качестве урны, возможно, заросли облепихи крушиновидной увеличивают интенсивность транспирации – как защитный механизм к негативным факторам окружающей антропосферы.
Содержание воды в листьях облепихи. Оводненность изучаемых листьев облепихи – один из важных признаков водного режима древостоев [11]. Как показано в табл. 3, содержание воды в пунктах Чолпон-Ата и Тамчы были высокими. Общая средняя величина содержания воды в листьях составила 45,31 %. Наименьший показатель среди исследуемых объектов показали образцы пункта Оттук, содержание воды в котором составило 32,15 %. В пунктах, где уровень антропогенного прессинга повышен, оводненность листьев была высокой (63,21 %), что, возможно, является адаптационным признаком к повышенной антропогенной нагрузке, но это требует дополнительных исследований и наблюдений.
Таблица 3
Средние значения содержания воды в изучаемых образцах
|
№ п/п |
Пункты отбора |
Cpeдниe знaчeния, М ± m, мг/г/чac |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
34,00 ± 0,18 |
1,32 |
|
2 |
Бостери |
34,00 ± 0,18 |
1,32 |
|
3 |
Чолпон-Ата |
63,21 ± 0,32 |
1,59 |
|
4 |
Тамчы |
63,21 ± 0,32 |
1,59 |
|
5 |
Оттук |
32,15 ± 0,16 |
1,25 |
Таблица 4
Водоудерживающая способность исследованных образцов
|
№ п/п |
Пункты отбора |
Cpeдниe знaчeния, М ± m, мг/г/чac |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
18,23 ± 0,41 |
9,58 |
|
2 |
Бостери |
19,16 ± 0,46 |
10,42 |
|
3 |
Чолпон-Ата |
33,53 ± 0,63 |
12,16 |
|
4 |
Тамчы |
29,14 ± 0,28 |
7,86 |
|
5 |
Оттук |
16,45 ± 0,45 |
6,32 |
Водопотеря. Показатели водопотери показали, что пункты, где повышенный уровень транспирации, понижена водоудерживающая способность листьев (табл. 4). Так, в пункте Чолпон-Ата среднее значение водопотери составило 33,53 мг/г/ч, близки к этому показателю данные по пункту Тамчы (29,14 мг/г/час), а наименьший показатель у пункта Оттук, где водопотеря составила 16,45мг/г/час. Таким образом, в наших исследованиях выявлена зависимость интенсивности транспирации с водоудерживающей способностью листьев, что было также отмечено и другими исследователями [12]. Возможно, повышенная нагрузка на экосистемы озера в пунктах Чолпон-Ата, Тамчы имеет негативное воздействие на дикорастущие заросли облепихи крушиновидной и требует разработки защитных мер по их сохранению.
Водопоглощение, или степень водного дефицита, показывает, сколько необходимо воды для восстановления водного дефицита. Наибольший показатель по водному дефициту был в образцах Чолпон-Ата (табл. 5), почти одинаковые результаты были в образцах Тамчы (15,32), Бостери (15,18), Булан-Соготту (14,05), а наименьший показатель водопоглощения был у образца Оттук (6,68). Таким образом, можно считать, что интенсивность транспирации оказывает влияние на степень водного дефицита растения.
Таблица 5
Средние показатели степени водного дефицита
|
№ п/п |
Пункты отбора |
Cpeдниe знaчeния, М ± m, мг/г/чac |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
16,05 ± 0,41 |
12,52 |
|
2 |
Бостери |
15,18 ± 0,46 |
13,18 |
|
3 |
Чолпон-Ата |
18,38 ± 0,63 |
7,26 |
|
4 |
Тамчы |
15,32 ± 0,28 |
13,45 |
|
5 |
Оттук |
6,68 ± 0,45 |
15,61 |
Таблица 6
Коэффициент засухоустойчивости
|
№ п/п |
Пункты отбора |
Cpeдниe знaчeния |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
1,01 |
7,27 |
|
2 |
Бостери |
1,04 |
8,14 |
|
3 |
Чолпон-Ата |
1,12 |
7,86 |
|
4 |
Тамчы |
1,09 |
8,51 |
|
5 |
Оттук |
1,1 |
6,45 |
Далее, согласно протоколу наших исследований, с целью определения комплексного коэффициента устойчивости, нами были определены коэффициенты засухоустойчивости, жаростойкости и солеустойчивости.
Коэффициент засухоустойчивости показывает степень выносливости древесного растения к водному дефициту. По литературным данным [13], чем ближе коэффициент засухоустойчивости к 1,0, тем более устойчивым к засухе видом принято считать растение. Наши исследования показали (табл. 6), что все изучаемые образцы обладали повышенной степенью устойчивости к водному дефициту и обладали повышенной засухоустойчивостью. Возможно, находясь длительное время в условиях повышенного антропогенного воздействия, дикорастущие заросли облепихи крушиновидной выработали повышенную степень засухоустойчивости, что очень важно в аспекте потепления климата и разработки адаптационных механизмов к изменению климата.
Коэффициент жароустойчивости. Наиболее высокий показатель по жароустойчивости изучаемых образцов был у образцов пункта Булан-Соготту (0,91), а наиболее низким – в пункте Чолпон-Ата (0,74), у остальных вариантов он варьировал в пределах 0,81 (табл. 7). Таким образом, можно судить о незначительном влиянии водного дефицита на жаростойкость образцов. Так в пункте Оттук, где наблюдался наименьший водный дефицит, коэффициент жаростойкости в среднем на 12,3 % выше чем в пунктах Булан-Соготту, Бостери и Тамчы, и на 23,0 % выше, чем в пункте Чолпон-Ата.
Таблица 7
Коэффициент жароустойчивости
|
№ п/п |
Пункты отбора |
Cpeдниe знaчeния |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
0,91 |
5,78 |
|
2 |
Бостери |
0,80 |
3,54 |
|
3 |
Чолпон-Ата |
0,74 |
3,68 |
|
4 |
Тамчы |
0,81 |
4,59 |
|
5 |
Оттук |
0,82 |
4,21 |
Таблица 8
Коэффициент солеустойчивости
|
№ п/п |
Пункты отбора |
Cpeдниe знaчeния |
Коэффициент вapиaции, Cv % |
|
1 |
Булан-Соготту |
1,51 |
9,65 |
|
2 |
Бостери |
1,47 |
8,52 |
|
3 |
Чолпон-Ата |
1,52 |
10,87 |
|
4 |
Тамчы |
1,40 |
8,14 |
|
5 |
Оттук |
1,80 |
7,49 |
Коэффициент солеустойчивости. Полученные данные о солеустойчивости изучаемых образцов (табл. 8) показали, что все образцы обладают повышенной солеустойчивостью, наибольшее значение коэффициента (1,80) было в пункте Оттук, наименьший – в пункте Тамчы (1,40), средний показатель был равен 1,54. Какой-либо зависимости между солеустойчивостью и ранее изученными показателями нами не было обнаружено.
По показателям трех вышеуказанных коэффициентов можно определить комплексный коэффициент устойчивости (ККУ) по отношению к таким экологическим факторам, как солеустойчивость, жаростойкость и засухоустойчивость. ККУ (табл. 9) был наиболее высоким в пункте Оттук (1,27), где сравнительно низкий рекреационный поток туристов, в остальных пунктах этот показатель менялся в сравнительно узких пределах (1,10–1,13 – в пункте Чолпон-Ата). Эти данные позволяют предполагать, что дикорастущие заросли облепихи крушиновидной выработали, хотя и незначительно, но адаптационный механизм к неблагоприятным факторам среды, что позволяет им расти и плодоносить в условиях повышенного антропогенного воздействия. Однако визуальные наблюдения за созревшими плодами показали, что большинство созревших плодов в пункте Чолпон-Ата (рис. 2) были загнившими.
Таблица 9
Показатели комплексного коэффициента устойчивости
|
№ |
Пункты отбора |
Коэффициeнты |
ККУ |
||
|
зacуxоуcтойчивоcти |
cолeуcтойчивоcти |
жapоуcтойчивоcти |
|||
|
1 |
Булан-Соготту |
1,01 |
1,51 |
0,82 |
1,11 |
|
2 |
Бостери |
1,04 |
1,47 |
0,80 |
1,10 |
|
3 |
Чолпон-Ата |
1,12 |
1,52 |
0,74 |
1,13 |
|
4 |
Тамчы |
1,09 |
1,40 |
0,81 |
1,10 |
|
5 |
Оттук |
1,1 |
1,80 |
0,91 |
1,27 |

Рис. 2. Созревшие плоды на побережье г. Чолпон-Ата (на фото Н. Тотубаевой отчетливо видно, что примерно половина плодов загнили уже на ветке)
Таким образом, можно предположить, что повышенное воздействие на экосистему озера Иссык-Куль может иметь негативное последствие, что было показано нами на примере дикорастущих зарослей облепихи крушиновидной, которое выражалось в увеличении интенсивности транспирации, приводящем к увеличению водного дефицита, что, к свою очередь, может привести к ослаблению растений и постепенному их изреживанию и вытеснению из ареала обитания. Как всем известно, в природе изменение и нарушение одной цепочки может привести к постепенным изменениям всей экосистемы, которое, в совокупности с всевозрастающим антропогенным воздействием на прибрежные экосистемы [14], может повлечь плачевные последствия для всей экосистемы и качества воды озера Иссык-Куль.
Библиографическая ссылка
Тотубаева Н.Э., Дуйшебекова С.Б., Кожобаев К.А. ВОДНЫЙ РЕЖИМ ЗАРОСЛЕЙ ОБЛЕПИХИ КРУШИНОВИДНОЙ (Hippophae rhamnoides) ПОБЕРЕЖЬЯ ОЗЕРА ИССЫК-КУЛЬ // Успехи современного естествознания. 2021. № 4. С. 77-83;URL: https://natural-sciences.ru/en/article/view?id=37611 (дата обращения: 07.07.2026).
DOI: https://doi.org/10.17513/use.37611