


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
ASSESSMENT OF BIOGENETIC DIVERSITY OF PLANTS OF FRUIT AND BERRY CROPS IN THE GENE POOL. CREATION OF NEW VARIETIES

В настоящей статье ставилась задача не ограничиваться феноизучением, темпами роста и развития образцов в генофонде, но изучить образцы с применением молекулярно-генетических методов. Это позволит дать цитогенетическую характеристику, выявить генотип, что особенно важно при создании новых сортов [1, 2]. Образцы земляники (Fragaria ananassa Duch) и унаби (Ziziphus jujube Mill.) в собранном генофонде Института генетических ресурсов НАН Азербайджана отличаются рядом свойств, величиной и качествами ягод и плодов, темпами роста и развития, биохимическими показателями [3–6].
Учитывая особенности климата республики с засушливым летом и наличием засоленных почв, отбор соле- и засухоустойчивых сортов земляники имеет большое значение [7–9]. Но только по фенопоказателям сортов невозможно выделить стрессоустойчивые формы. Использование цитогенетического анализа, содержания ДНК, структуры ДНК хроматина, показателя хромосомного комплекса выявило проявление генотипа. Анализы генотипов и фенотипов различных образцов земляники помогут выявить разницу между стрессоустойчивыми и стрессонеустойчивыми сортами [9, 10]. Эти показатели можно рассматривать как генетические маркеры по выявлению стрессустойчивых сортов земляники в определенных экологических зонах.
Унаби растет в республике издавна, но местные сорта и формы в основном суховатые, невысокого вкуса. Последнее время с использованием ряда старых методик в сочетании с генетико-цитологическими исследованиями получены новые, крупноплодные сорта и гибриды [11].
Изучение фено- и генетикоцитологического разнообразия образцов культур: земляники и унаби, отбор новых форм. Применение в селекционных работах наряду с традиционными методами цитогенетических анализов, которые в последнее время приобретают большое значение в ряде областей биологии, своевременно.
Материалы и методы исследования
Работа проводилась на Апшеронской опытной базе Института генетических ресурсов НАН Азербайджана. Работа заключалась в изучении фенотипа и генотипа образцов земляники и унаби в генофонде, в проведении цитогенетических исследований, особенностей хромосомного комплекса.
Контроль генетической устойчивости к солевому стрессу (0,2 М NaCl) и осмотическому стрессу (ПЕГ-0,5 атм) заключался в изучении восстановительных особенностей земляники, после использования стресс-агентов. Исследования включали данные по биосинтезу ДНК и структуры ДНК хроматина у различных сортов. Эти данные показатели можно рассматривать как генетические маркеры по выявлению стрессоустойчивых сортов земляники [8, 9]. Параллельно с растениями земляники наряду с феноизучением сортов и форм велись с унаби также исследования по содержанию ДНК, структуры ДНК хроматина, его упаковки, жаровыносливости.
Содержание ДНК определяли спектрофотометрически при длине волны 270 и 290 мм.
Метод фракционирования ДНК проводился по принципу ступенчатого воздействия на хроматин разной ионной силы, что позволяет разделить клеточную ДНК на лабильную, стабильную и остаточную фракции. Определение содержания отдельных фракций ДНК проводилось на свежем листовом материале земляники и унаби методом центрифугирования (500–2000 об.) и дальнейшим спектрофотометрированием. Образцы просматривались на спектрофотометре.
Характер наследования признаков гибридами определялись по соотношению среднего показателя гибрида к среднему показателю родителей по формуле
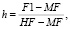
где h – степень наследования; F1 – среднее значение признаков гибридного поколения; HF – значение признаков лучшего родителя; MF – среднее значение признаков обоих родителей.
Цитологические анализы земляники и унаби проводились по общепринятой для плодовых культур методике [12].
Результаты исследования и их обсуждение
У неустойчивых к стрессам сортам земляники Дильбер и Гибрид 237 отмечено ослабление синтеза ДНК на 45 %. У стрессоустойчивых сортов Берюлевская, Лермонтовская, Кавальер, Реддекоут по содержанию ДНК, отмечено увеличение этого показателя почти на 50 %. У высоко- и среднеустойчивых сортов отмечено увеличение остаточной прочной связанной фракции ДНК и почти трехкратное уменьшение этого показателя у стрессонеустойчивых сортов (табл. 1). А остаточная фракция редко дает такие явные результаты. Проявление генотипа у некоторых сортов земляники позволяет выявить варьирования локусов изменчивости признаков.
Таблица 1
Содержание и cтруктурно-функциональное состояние ДНК хроматина в зависимости от стрессоустойчивости у сортов и форм крупноплодной земляники
|
№ |
Сорта и формы |
ДНК / фракции мг/100 г |
Устойчивость сортов к стрессам |
||||
|
лабильная |
стабильная |
остаточная |
общее |
% к контролю |
|||
|
1 |
Лермонтовская |
0,64 |
0,72 |
1,59 |
2,95 |
─ |
Высокоустойчивый |
|
NaCE |
1,46 |
1,42 |
3,02 |
5,90 |
+75,9 |
||
|
PEQ |
0,76 |
1,18 |
2,45 |
4,40 |
+49,1 |
||
|
2 |
Кавальер |
2,88 |
2,19 |
1,62 |
6,69 |
─ |
Высокоустойчивый |
|
NaCE |
3,43 |
3,52 |
2,65 |
9,60 |
+43,4 |
||
|
PEQ |
3,85 |
4,73 |
2,76 |
11,34 |
+69,5 |
||
|
3 |
Берюлевская |
1,66 |
0,79 |
5,58 |
8,03 |
─ |
Среднеустойчивый |
|
NaCE |
1,97 |
1,64 |
6,09 |
9,70 |
+22 |
||
|
PEQ |
2,39 |
1,09 |
6,87 |
10,35 |
+29 |
||
|
4 |
Реддекоут |
2,46 |
2,93 |
1,01 |
6,39 |
─ |
Среднеустойчивый |
|
NaCE |
2,53 |
3,77 |
1,33 |
7,63 |
+19,4 |
||
|
PEQ |
1,28 |
1,06 |
0,58 |
7,63 |
-55 |
Засухонеустойч. |
|
|
5 |
Дильбер |
9,09 |
2,14 |
1,79 |
13,02 |
─ |
неустойчивый |
|
NaCE |
8,51 |
2,87 |
1,19 |
12,57 |
-4 |
||
|
PEQ |
8,15 |
2,05 |
1,16 |
11,36 |
-12,7 |
||
|
6 |
Гибрид 237 |
6,40 |
2,07 |
1,06 |
9,53 |
─ |
неустойчивый |
|
NaCE |
4,18 |
2,20 |
0,66 |
7,04 |
-26,5 |
||
|
PEQ |
3,59 |
1,33 |
0,33 |
5,25 |
-45,0 |
||
Образцы генофондной коллекции унаби также различаются фенотипами величиной плодов, темпами роста и развития, генотипом.
Исследованиями установлено, что местные сорта и формы унаби представляют интерес при создании гибридов в качестве источника устойчивости к болезням и вредителям, засухоустойчивости, интродуценты являются донорами крупноплодности, высоких диетических качеств [13]. Результат анализов 200 гибридных саженцев унаби, полученных гибридизацией между растениями местными мелкоплодными♀ и крупноплодными интродуцентами♂, свидетельствуют о сложном характере наследования величины плодов. Данные результаты объясняются гетерозиготностью родительских компонентов и полигенным характером изучаемых признаков (табл. 2). Примерно у 50 % полученных гибридных образцов отмечен промежуточный характер наследования величины плодов между местными мелкоплодными и крупноплодными интродуцентами, степень доминирования h = 0, эти показатели колеблются 0,11–0,18. У 33–37 % гибридов отмечено частичное или полное доминирование мелкоплодной материнской формы, где h колебалось от -0,1 до -0,54 и 15–17 % новых гибридных сеянцев имело h (доминирование отцовских крупноплодных сортов в интервале от + 0.55 до + 0.18). В одних и тех же комбинациях скрещивания отмечено расщепление гибридов по величине, массе плодов на промежуточное наследование, наследование отцовского или материнского показателя, т.е. 3 степени (h = 0, h > 0, h < 0).
Таблица 2
Характеристика наследования массы плодов у гибридов унаби
|
N |
Гибриды |
Комбинация скрещивания (сорта) |
М ± м (г) |
h степень домини рования |
Характер наследования |
||
|
Мелко плодный ♀ |
Гибрид F |
Крупно плодный ♂ |
|||||
|
1 |
1/4 |
♀мелкоплодХ ♂Таян-цзао |
3,5 ± 0,12 |
8,6 ± 0,27 |
19,0 ± 0,46 |
-0,37 |
Част. домин. Мелкоплод. ро-ля |
|
2 |
6/18 |
♀мелкоплодХ ♂Таян-цзао |
3,5 ± 0,12 |
12 ± 0,47 |
19,0 ± 0,46 |
0,18 |
Промежуточное |
|
3 |
9/1 |
♀мелкоплодХ ♂Таян-цзао |
3,5 ± 0,12 |
10,3 ± 0,36 |
19,0 ± 0,46 |
-0,12 |
Промежуточное |
|
4 |
9/2 |
♀мелкоплодХ ♂Таян-цзао |
3,5 ± 0,12 |
7,0 ± 0,27 |
19,0 ± 0,46 |
-0,54 |
Домин. мелкоп. р-ля |
|
5 |
13/1 |
♀мелкоплодХ ♂Таян-цзао |
3,5 ± 0,12 |
8,5 ± 0,46 |
19,0 ± 0,46 |
-0,35 |
Домин. мелкоп. р-ля |
|
6 |
3/2 |
♀мелкоплодХ ♂Юбилейный |
3,5 ± 0,12 |
7,0 ± 0,69 |
8,0 ± 0,31 |
+0,55 |
Домин. круп. р-ля |
|
7 |
4/8 |
♀мелкоплодХ ♂Юбилейный |
3,5 ± 0,12 |
6,3 ± 0,31 |
8,0 ± 0,51 |
0,24 |
Промежуточное |
|
8 |
3/2 |
♀мелкоплодХ ♂Юбилейный |
3,5 ± 0,12 |
8,0 ± 0,21 |
8,0 ± 0,51 |
+1,40 |
Домин. круп. р-ля |
|
9 |
2/1 |
♀мелкоплодХ ♂Даргомский |
3,5 ± 0,12 |
8,9 ± 0,21 |
17,5 ± 0,51 |
-0,22 |
Част. домин. Мелкоплод. ро-ля |
|
10 |
15/1 |
♀мелкоплодХ ♂Даргомский |
3,5 ± 0,12 |
11,3 ± 0,16 |
17,5 ± 0,51 |
0,11 |
Промежуточное |
Исследование у унаби метаболизма ДНК по образцам в генофонде велось в двух направлениях:
1) исследование содержания ДНК у гибридов унаби, структуры ДНК хроматина, его упаковки;
2) изучение генома унаби у стародавних и новых селекционных сортов в коллекции.
По первому направлению работы получены свидетельства усиления механизма редупликации ДНК и образования копий генов у гибридных растений при частичном или полном доминировании величины плодов одной из родительских форм [1].
При промежуточном наследовании величины плодов у гибридов часто содержание ДНК, лабильной фракции ДНК хроматина занимало промежуточное положение. При доминировании величины плодов одной из родительских форм процесс метаболизма нуклеиновых кислот идет в основном как за счет увеличения лабильной фракции ДНК, так и более высокого его долевого содержания.
По второму направлению получены данные, свидетельствующие, что у местных стародавних сортов и стародавнего сорта Таян-цзао содержание ДНК в 1,5–2 раза ниже, чем у новых средних крупноплодных сортов, часто гибридного происхождения. Предполагается наличие механизмов регуляции ДНК за счет сброса повторов в геноме. У новых часто гибридных сортов содержание ДНК в клетке приводит к образованию копий генов или повторностей. У старых сортов показатель изменчивости имеет малый разброс 1,6–2,1 %, у новых изменчивость геномов имеет высокую амплитуду колебания от 0,7 % у нового сорта Даргомский, до 12,6 % еще более молодого сорта Вахш.
Как видно из рисунка, местные сорта характеризуются и меньшим количеством ДНК на клетку. Эти данные говорят о нестабильности генома новых сортов, их общей пластичности, изменчивости по сравнению со старыми. По содержанию ДНК в клетках стародавние сорта уступают новым сортам. Аналогичный процесс наблюдается по структурно-функциональному состоянию ДНК хроматина. Содержание лабильной фракции ДНК хроматина колеблется у стародавних сортов (-0,062 пг) в клетке, у новых сортов этот показатель характеризуется высокой амплитудой колебания с разницей (+0,277 пг) у новых сортов.
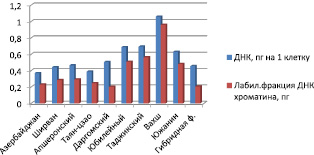
Синтез ДНК и лабильной фракции ДНК в клетках стародавних и новых сортов растений зизифуса
Показатель активности генетического аппарата новых сортов характеризуется высоким разбросом в сравнении со стародавними сортами.
Исследования хромосомного комплекса различных образцов, как земляники, так и унаби свидетельствуют о полиморфизме этого показателя. Коллекции и земляники, и унаби включали основное число диплоидных сортов (2n = 24) унаби, триплоидные (2n = 36), у нового сорта Даргомский наряду с 24 хромосомами отмечено появление 2n = 12, 28, 30, 36 (1 клетка) и 2 клетки 2n = 48. Аналогичная картина полиморфизма этого показателя 2n отмечена у земляники основное число 2n = 42 (гексаплоид), обнаружены образцы с клетками 2n = 14, 28, 56 этих культур.
Исследованиями выявлено фено- и цитогенетическое разнообразие (полиморфизм) образцов генофондов земляники и унаби.
Модификация структуры хроматина происходит за счет распределения фракций хроматина, их содержания, доли, особенно лабильной фракции, являющейся маркером сортов, у гибридов земляники при ее регуляции засухо- и солеустойчивости [7, 11, 14].
Были изучены различные сорта и формы земляники и унаби, что свидетельствовало о существенных различиях, обусловленных генетическими особенностями образцов. Выявлена связь активизации ДНК со стрессустойчивостью, увеличение лабильной фракции, эухроматизации хроматина у стрессоустойчивых сортов земляники. Это положение актуально для республики с ее высокими летними температурами, засухами, наличием засоленных земель. Отмечены модификации ДНК хроматина у стрессоустойчивых и неустойчивых сортов земляники: резкое повышение содержания лабильной фракции у первых и снижение этого показателя у вторых.
Используя данные биоцитогенетических анализов, авторы создали новые, перспективные сорта, адаптированные к местным условиям, изучены стрессоустойчивые сорта земляники, которые могут заменить стрессонеустойчивые сорта. Используя различные методы биологии, соавторы пополнили генофонд новыми сортами, которые заменили старые, стрессонеустойчивые. Также авторами статьи выведены два новых, крупноплодных сорта унаби, переданные в Госсорткомиссию.
Заключение
1. Генофонд земляники и унаби состоит из многочисленных образцов, имеющих разный фенотип, генотип, различные темпы роста и развития.
2. Коллекция земляники и унаби полиморфна по содержанию ДНК, лабильной фракции ДНК хроматина, по комплексу хромосом. У одного сорта встречаются ди-, три-, тетра-, гексаплоидные экземпляры. Чаще всего встречаются основные числа у земляники 2n = 42 (гексаплоид), у унаби основное число 2n = 24 (диплоид).
3. Степень доминирования величины плодов (масса, длина, диаметр) в коллекции очень разнообразна. При гибридизации крупноплодных и мелкоплодных плодов гибриды характеризуются в основном промежуточным доминированием (h = 0), но отмечено частичное появление гибридов, схожих с мелкоплодным материнским (h < 0) и реже с крупноплодным отцовским (h > 0) типом. В одной комбинации скрещивания часто возникают три группы гибридов.
4. Установлено изменение нуклеинового метаболизма (содержание ДНК, структуры ДНК хроматина, упаковки хроматина) у стрессоустойчивых образцов земляники и гибридных форм унаби с различным генотипом стародавних и новых сортов. Очевидно, существуют механизмы регуляции, выражающиеся в сбросе повторов.
5. Выделены и отобраны различными путями (отбор, гибридизация, по итогам изучения структурно-функционального состояния ДНК, упаковки хроматина у обеих культур), новые формы с рядом положительных признаков, адаптированные к местным условиям.
Библиографическая ссылка
Гаджиева А.Ф., Ахундова Н.И., Гасанов Н.А. ОЦЕНКА БИОГЕНЕТИЧЕСКОГО РАЗНООБРАЗИЯ РАСТЕНИЙ ПЛОДОВО-ЯГОДНЫХ КУЛЬТУР В ГЕНОФОНДЕ. СОЗДАНИЕ НОВЫХ СОРТОВ // Успехи современного естествознания. 2021. № 6. С. 7-12;URL: https://natural-sciences.ru/en/article/view?id=37634 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/use.37634