


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
BIOLOGICAL ACTIVITY OF FROZEN SOILS

Микробная биомасса, входящая в состав органического вещества почвы, является важным звеном в цикле углерода и может быть использована в качестве показателя биологической активности почвы. Общие резервуары углерода микробной биомассы имеют невысокие значения, как правило, они составляют не более 10% от общего содержания углерода почвы, только в некоторых случаях могут достигать 15% [1]. Микробная биомасса играет важную роль в минерализации отмершего растительного опада, трансформации ряда элементов – С, N, P, S, иммобилизации тяжелых металлов и сохранении почвенной структуры [2].
Микроорганизмы являются важным компонентом экологической функций почв. По ряду физических и химических характеристик микроорганизмы являются ключевой частью поддержания жизнедеятельности равновесных экосистем. Это достигается вследствие высоких значений биомассы и благодаря их таксономическому разнообразию. Почвы всегда имеют запасной пул для обеспечения большей устойчивости, состоящий из разных микроорганизмов, представленный: наноформами, спорами, некультивируемыми и латентными формами. При нарушении саморегуляции системы они становятся функциональными [2].
В превращении всех биогенных макроэлементов экосистемы микробная биомасса играет исключительно важную роль. Деструкция и минерализация органического вещества почвы и поступление углерода в биосферу происходит благодаря почвенным микроорганизмам. Для определения состояния почв можно использовать показатель микробной биомассы [3]. Она является одним из индикаторов изменения биологической активности почв, характеризуя степень биогенности почвы. На основе количественной оценки микробной массы проводится оценка круговорота углерода в экосистемах и биосфере.
Потенциал микробного пула в способности почвы сохранять нормальное функционирование микробных сообществ под воздействием неблагоприятных климатических и антропогенных воздействий является важным эколого-физическим показателем [2; 4; 5]. Информация о пулах микробного углерода в разных типах почв важна для прогнозных сценариев и для экологических исследований.
Понимание фундаментальных основ этих процессов становится особенно важным с учетом глобальных изменений природной среды и климата, которые происходят с деятельностью человека. При исследовании запасов углерода микробной биомассы в разных типах почв, количество его накопления является основным показателем в исследованиях.
Биологическая активность почв является индикатором экологического состояния природных экосистем. Изменения в биологической активности почвы могут неизбежно оказывают влияние на круговорот веществ. Биологическая активность почв является одной из важных характеристик интенсивности микробиологических процессов. В качестве характеристики биологической активности нами использован показатель углерода микробной биомассы.
Биологическая активность черноземов квазиглеевых, черноземов глееватых криотурбированных и гумусово-квазиглеевых окарбоначенных почв юга Витимского плоскогорья изучены недостаточно полно, поэтому нуждаются в дополнительных исследованиях.
Цель работы – провести мониторинг накопления углерода микробной биомассы, а также определить его запасы в мерзлотных почвах для оценки общей биологической активности мерзлотных почв.
Материалы и методы исследования
Экспериментальные участки находятся на юге Витимского плоскогорья, Еравнинская котловина. Многолетняя мерзлота достигает 130 метров, средний показатель равен 85 метрам [6]. На глубине 1,5–3,0 метра от поверхности почвы залегает верхняя граница мерзлоты.
Еравнинская котловина характеризуется резко континентальным климатом с холодной, малоснежной и продолжительной зимой. Последняя сменяется поздней засушливой ветреной весной. Лето в районе исследования жаркое и короткое. Данные метеостанции с. Сосново-Озерское свидетельствуют о том, что среднегодовая температура воздуха составляет – 4,1 °C при средней температуре наиболее теплого месяца (июль) +17,1 °C и самого холодного (январь) -25,4 °C. Сумма биологически активных температур составляет 1330 °С.
Состав растительного покрова участков исследования многообразен. Встречаются злаково- и разнотравно-пижмовые, вострецовые и вострецово-ковыльные растительные ассоциации. На более влажных экотопах произрастают овсяницы, тонконог стройный, ковыльный волосатик. При учете надземной, подземной и общей фитомассы было выявлено, что биопродуктивность растительной биомассы черноземов глеевых криотурбированных составляет 2,08 кг/м2 и находится наравне с черноземами квазиглеевыми – 2,05, что значительно ниже по сравнению с таковыми гумусово-квазиглеевыми окарбоначенными почвами – 2,50 кг/м2. Почвообразующие породы: делювиально-карбонатные суглинистые отложения, слоистые озерные отложения.
Объектами исследования послужили черноземы квазиглеевые, черноземы глееватые криотурбированные и гумусово-квазиглеевые окарбоначенные почвы. Для проведения исследования выбраны экспериментальные площадки, где заложены разрезы, взяты образцы почв с шагом 10 см. Мощность сезонно-талого слоя для черноземов квазиглеевых составляет 285 см. Данные по микробиоценозу мы приводили в 0-20 см слое почвы.
Биологическая активность оценивалась по интенсивности накопления, динамике и запасам углерода микробной биомассы в почвах.
Классическим методом определения углерода микробной биомассы является регидратационный метод [7]. Метод основывается на высушивании почвы при температуре 65-70 °С, что ведет к деструкции цитоплазматической мембраны микроорганизмов. На органическое вещество, которое уже отмерло, фактор температуры практически не воздействует. Внутриклеточные компоненты микроорганизмов благодаря регидратации высвобождаются и переходят в раствор слабой нейтральной соли и в дальнейшем определяются по сумме органических соединений. В воздушно-сухих образцах углерод микробной биомассы имеет некорректные заниженные данные, поэтому анализировать нужно только свежие образцы почвы. Почвенные образцы отбирались при n=3 и глубине 0-20 см.
С подробной характеристикой разрезов можно ознакомиться в статье [8].
Основные характеристики черноземов квазиглеевых: мощность гумусового горизонта – небольшая, гранулометрический состав – средний суглинок, содержание углерода органического в верхнем горизонте – среднее, сумма поглощенных – высокая, реакция среды в верхней части профиля – слабокислая (рН=6,52) и близкая к нейтральной (рН=7,15), в нижней части – щелочная реакция (рН=8,36).
Морфологические характеристики исследованных черноземов глееватых криотурбированных и гумусово-квазиглеевых окарбоначенных почв значительно отличаются от черноземов квазиглеевых, где первые в значительной степени затронуты криогенными процессами на уровне микрорельефа. Реакция среды гумусово-квазиглеевых окарбоначенных почв – щелочная (8,02–8,40), черноземов глееватых криотурбированных – слабощелочная (7,71-7,81). По гранулометрическому составу черноземы глееватые криотурбированные относятся к супесчаным и легкосуглинистым, гумусово-квазиглеевые окарбоначенные почвы – легкосуглинистые в 0–30 см слое, среднесуглинистые в слое 30–50 см. Содержание органического углерода (Сорг) в черноземах глееватых криотурбированных составило 11,11% в 0-10 см слое, 6,38% – в 10-20 см слое. Отмечено высокое содержание Сорг в гумусово-квазиглеевых окарбоначенных почвах в слое 0–10 см – 17,56%, в слое 10-20 см – 15,85% (n=3).
Результаты исследования и их обсуждение
Микробиоценоз криогенных экосистем формируется под воздействием экстраконтинентальных природных условий. Изменения экологических условий способствуют формированию адаптационной реакции у микроорганизмов, поскольку суровый климат мерзлотной лесостепи повышает их уязвимость.
Энергетические и трофические факторы лимитируют накопление углерода микробной биомассы в период вегетации, при большом интервале температур и влажности. На накопление микроорганизмов и функциональную ее активность влияют экстремальные природные условия. Например, после долгой засухи при выпадении осадков мы можем наблюдать влияние влажности. При промерзании-оттаивании почв выражено влияние температуры. Микробная биомасса может в несколько раз уменьшаться, например при отмирании во время засухи, или увеличиваться при оптимизации гидротермических условий почв [9].
Сукцессия микроорганизмов в изменяющихся условиях среды определяет процесс роста микробной биомассы. За источники питания усиливается конкуренция между растительными и микробными сообществами. На замедление роста содержания микроорганизмов влияют изменения трофических факторов и повышение температуры почвы в сочетании с недостатком влаги. Благодаря активному поступлению корневых выделений складываются благоприятные условия для роста и развития r-стратегов только в ризосфере и ризоплане. Активность роста ризосферных микроорганизмов практически не оказывает влияния на общее накопление углерода микробной биомассы в почвах, что обусловлено низкими границами микрозон, не превышающими 250-1000 мкм. При таких условиях возможно доминирование К-стратегов, которые более устойчивые к неблагоприятным факторам среды и способны быстро расти [9].
Среднее накопление углерода микробной биомассы в верхнем 0-20 см слое черноземов глееватых криотурбированных равен 44,2 мг/100 г почвы (рис. 1).
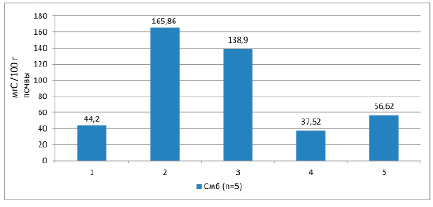
Рис. 1. Накопление углерода микробной биомассы в холодных и мерзлотных почвах. Условные обозначения: 1 – чернозем глееватый криотурбированный, 2 – гумусово-квазиглеевая окарбоначенная почва, 3 – чернозем квазиглеевый, 4 – каштановая почва [10], 5 – чернозем дисперсно-карбонатный [11]
Максимальное количество углерода микробной массы обнаружено в гумусово-квазиглеевой окарбоначенной почве – 165,86 мг/100 г почвы, что объясняется обилием ризосферной микрофлоры и высоким содержанием органического углерода. В черноземах квазиглеевых среднее значение углерода микробной биомассы в верхнем слое почвы 138,9 мг/100 г почвы. При усилении степени аридизации показатель уменьшается до 37,52 в каштановых почвах [10; 11].
Накопление углерода микробной биомассы в черноземе глееватом криотурбированном меньше, несмотря на то что в нем больше углерода, чем в черноземах квазиглеевых. Вероятно, этот углерод представлен большей частью недоступной для микробиоты формой.
Следует отметить, что видовая структура и разнообразие почвенных микроорганизмов может обедняться вследствие использования почвенными микробами легких органических соединений, которые содержатся в растительных остатках и корнях растений. Химический состав ризовыделений различен у растительности разных типов землепользования из-за различий в видовом составе растений. Корневой углерод является основным ресурсом питания микробных сообществ. Высвобождение органических соединений через корни (ризоотложения) и последующее их использование микробными сообществами является ключевым процессом, связывающим атмосферный и почвенный углерод и круговорот питательных веществ [12].
Динамика углерода микробной биомассы имеет одновершинный характер с максимумом в летний период. Так, в начале вегетационного сезона показатель варьирует от 40,3 мг С/100 г почвы в черноземах глееватых криотурбированных до 173,0 мг С/100 г почвы в гумусово-квазиглеевых окарбоначенных почвах, в черноземах квазиглеевых до 148,8 мг С/100 г почвы.
Летнее увеличение количества углерода микробной биомассы может быть связано с благоприятными гидротермическими условиями и бурным ростом r-стратегов [9]. Показатель варьирует от 78,8 мг С/100 г почвы в черноземах глееватых криотурбированных до 247,7 мг С/100 г почвы в гумусово-квазиглеевых окарбоначенных почвах, черноземах квазиглеевых – 168,8 мг С/100 г почвы (рис. 2).
Спад содержания углерода микробной биомассы в конце вегетации связан с торможением биологических процессов в почве в связи с понижением внутрисуточных температур почвы. Было отмечено понижение показателей углерода микробной биомассы до 48,8 в черноземах глееватых криотурбированных, до 204,5 мг С/100 г почвы в гумусово-квазиглеевых окарбоначенных почвах, до 143,0 мг С/100 г почвы в черноземах квазиглеевых – 148,8 мг С/100 г почвы.
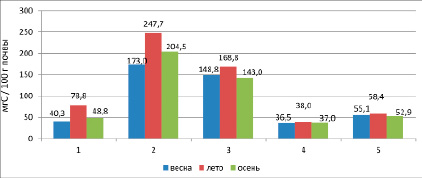
Рис. 2. Динамика углерода микробной биомассы в холодных и мерзлотных почвах. Условные обозначения: 1 – чернозем глееватый криотурбированный, 2 – гумусово-квазиглеевая окарбоначенная почва, 3 – чернозем квазиглеевый, 4 – каштановая почва [10], 5 – чернозем дисперсно-карбонатный [11]
Для сравнения приведем данные не с мерзлотных почв. Так, в каштановых почвах и черноземах дисперсно-карбонатных Тугнуйской котловины [10] динамика углерода микробной биомассы количественно меньше исследуемых мерзлотных почв. Кривая динамики углерода микробной биомассы имеет пологий одновершинный характер (рис. 2).
Изменения накопления углерода микробной биомассы связаны с воздействием трофических и климатологических факторов. Особенности суточных колебаний роста микроорганизмов в разных типах почвы показывают наличие единого эндогенного и экзогенного устройства сукцессионных сдвигов микробного сообщества.
Расчёты статистических параметров показали, что на различных типах почв накопление углерода микробной биомассы достоверно отличается. Установлена достоверная связь между температурой почвы и углеродом микробной биомассы в почвах. В отдельные периоды вегетации дефицит влажности в почвах сдерживал рост микроорганизмов. Корреляционная связь между углеродом микробной биомассы и влажностью почвы составляет 0,6-0,9.
Средние значения углерода микробной биомассы за весь период мониторинга имеют вариацию в широком диапазоне, что показывает способность микроорганизмов откликаться на изменение окружающей среды.
Запасы углерода микробной биомассы распределены в следующем ряду по убыванию: черноземы квазиглеевые – 0,22 кг С/м2, гумусово-квазиглеевые окарбоначенные почвы – 0,18 кг С/м2, черноземы глееватые криотурбированные – 0,08 кг С/м2 (рис. 3). Для сравнения в немерзлотных почвах: 0,07 кг С/м2 в каштановых [10] и 0,13 кг С/м2 в черноземах дисперсно-карбонатных [11].
Значительное накопление углерода микробной биомассы в условиях в засушливые периоды объясняется адаптационной реакцией микроорганизмов к условиям окружающей среды. По-видимому, в этом случае температура для их существования не играет значительной роли. Отмечено, что при очень низких значениях влажности почвы могут функционировать такие микроорганизмы, как ряд грибов и актиномицеты. Таким образом, внутри пула органического углерода микроорганизмами осуществляется трансформация углерода из углерода гумуса в углерод биомассы.
Абиотические условия каждого сезона определяют значительную трансформацию структуры микробиоценоза, пропорции отдельных микроорганизмов и их количества. Климатические условия в течение вегетационных периодов определяют различия в численности микроорганизмов. Условия формирования микроорганизмов обусловлены их физическими качествами и определяют сукцессию сообщества микроорганизмов. Периоды всплесков накопления микроорганизмов обусловлены абиотическими факторами (влажность, температура, осадки). При этом ограничительным фактором является низкий процент почвенного гумуса.
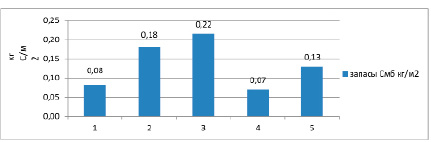
Рис. 3. Запасы углерода микробной биомассы в холодных и мерзлотных почвах. Условные обозначения: 1 – чернозем глееватый криотурбированный, 2 – гумусово-квазиглеевая окарбоначенная почва, 3 – чернозем квазиглеевый, 4 – каштановая почва [10], 5 – чернозем дисперсно-карбонатный [11]
Отличия в источнике питания и интенсивности влияния на почвы мерзлоты обусловливают разный запас питания, отсюда и различия содержания и динамики углерода биомассы.
Заключение
В ходе исследования достоверно определено, что биологическая активность гумусово-квазиглеевой окарбоначенной почвы выше, чем чернозема глееватого криотурбированного. Все изученные почвы отличаются по содержанию микробной биомассы. Невысокий уровень энергетического обеспечения, глубокое промерзание почвенного профиля и значительное их иссушение в весенне-летний период отразились на почвенной биоте черноземов глееватых криотурбированных.
Изменения содержания углерода микробной биомассы в течение вегетационного периода определяются гидротермическими условиями почв и общим ресурсом энергии. Углерод микробной биомассы исследуемых почв занимает невысокую процентную долю в общем пуле органического углерода. Несмотря на низкое процентное содержание в общем пуле органического углерода, микробная биомасса представляет собой высокофункциональную ее часть, способную значительно усилить темп круговорота почвенного органического вещества.
Экспедиционные исследования выполнены за счет средств бюджета по теме госзадания № 121030100228–4; аналитические работы при финансовой поддержке в рамках гранта РФФИ №16–04–01297.
Библиографическая ссылка
Чимитдоржиева Э.О., Корсунова Ц.Д-Ц., Чимитдоржиева Г.Д. БИОЛОГИЧЕСКАЯ АКТИВНОСТЬ МЕРЗЛОТНЫХ ПОЧВ // Успехи современного естествознания. 2022. № 12. С. 206-211;URL: https://natural-sciences.ru/en/article/view?id=37972 (дата обращения: 02.07.2026).
DOI: https://doi.org/10.17513/use.37972