



В настоящее время проблему, связанную с процессом роста, размножения и гибели микроорганизмов, целесообразно разделить на микрокинетическую и макрокинетическую. Микрокинетическая модель должна изучать рост, способ размножения и гибель отдельной изолированной клетки в зависимости от ее физиологического состояния, компонентов субстрата в ближайшем окружении клетки и скорости метаболизма в целях определения вероятности элементарных актов процесса роста, деления и гибели микроорганизма за единицу времени в заданном объеме системы.
В круг задач макроскопической модели следует включать вывод, исследование и разработку математических методов решения кинетических уравнений, описывающих эволюцию функции распределения популяции по массам (объемам) при заданном законе роста, размножения и гибели отдельной клетки.
Уравнения микрокинетики роста клетки имеют следующий вид:
![]()
- когда процесс микробиологического синтеза лимитируется биохимическими превращениями внутри клетки (Варфоломеев, Гуревич, 1999);
![]()
- если процесс микробиологического синтеза лимитируется диффузионным переносом питательных веществ к поверхности клетки.
Здесь
m0 ≤ m(t) ≤ 2m0, 0 ≤ t ≤ tg,
![]() ψ(t) = ψ (C),
ψ(t) = ψ (C),
m(t) - масса отдельной клетки в момент времени t (m - детерминированная величина); t = tg - время генерации (деления клетки); m0 - масса клетки в момент времени t = 0; U(m, С) - скорость роста клетки; С - концентрация субстрата; µ0 и Кс - постоянные величины; β - коэффициент массообмена; ![]() где DM - коэффициент молекулярной диффузии, зависящий от температуры культуральной жидкости, d - толщина «пограничной пленки», зависящая от гидродинамической обстановки в окрестности клетки; S(t) - площадь внешней поверхности клетки в момент времени t. Предполагается, что при m(t) = 2m0 происходит деление клетки на две дочерние.
где DM - коэффициент молекулярной диффузии, зависящий от температуры культуральной жидкости, d - толщина «пограничной пленки», зависящая от гидродинамической обстановки в окрестности клетки; S(t) - площадь внешней поверхности клетки в момент времени t. Предполагается, что при m(t) = 2m0 происходит деление клетки на две дочерние.
Синхронное деление клеток. Согласно Дж. Бейли и Д. Оллису (1989) на ограниченном отрезке времени можно создать такие условия в системе, когда все клетки будут делиться синхронно. В этом случае при ![]() имеют место следующие закономерности:
имеют место следующие закономерности:
μ(t) =m02 τ, τ < 1;
![]() N(t) = N02 E(τ) , M(t) = M02τ,
N(t) = N02 E(τ) , M(t) = M02τ,
τ ≥ 0,
1. где M(t) и N(t) - масса и число клеток микроорганизмов в момент времени t>0 в единице объема системы M0 = M(0), N0 =N(0), Е(τ ) - целая часть от τ, равная наибольшему целому числу, не превосходящему τ. Так, Е(τ ) = 0 при 0<τ <1; Е(τ ) = 1 при τ = 1 + 0 и Е(τ ) = 0 при τ = l -0). Очевидно, что M(t)/N(t) = x(t) = m02 τ -Е(τ ) - периодическая функция t с периодом Т = 1. Однако если, например, материнская клетка растет по закону ![]() , где u0 - постоянная величина, то
, где u0 - постоянная величина, то
m(t) = m0(1+τ1), τ1<1,
τ1=t/τ0 τ0=m0/u0, ![]() ,
,
x(t) = m0[1+τ1 - E(τ1)], N(t) = N02 E(τ),
M(t) = x(t)N(t), τ >0.
2. По мере роста популяции ступенчатая зависимость числа клеток нарушается, что обусловлено стохастической природой роста и размножения микроорганизмов. Более того, как правило, популяция микроорганизмов состоит из громадного числа клеток, асинхронно делящихся и растущих с некоторыми индивидуальными скоростями в соответствии с их возрастом, со случайным изменением концентрации компонентов и метаболизма в ближайшем окружении отдельной клетки, каждая из которых представляет собой миниатюрный биореактор переменного объема, в котором непрерывно перерабатываются питательные вещества и синтезируются новые необходимые для жизнедеятельности микроорганизма.
Уравнения макрокинетики. При построении математической модели рассматриваемого процесса проблемами первоочередной важности являются выявление общих закономерностей и разработка математических методов описания эволюционных процессов, связанных с ростом, размножением и гибелью микроорганизмов в пространственно-однородных и неоднородных дисперсных системах, на основании единого логического подхода. Но в общем случае математическое описание процесса формирования спектра масс микроорганизмов в дисперсной среде чрезвычайно сложная задача. Однако если столь сложный процесс рассматривать как совокупность таких менее сложных («элементарных») процессов, участвующих в формировании спектра масс микроорганизмов, как непрерывный рост, размножение клеток, гидродинамическая обстановка в системе и др., то для математического описания сложного процесса целесообразно использовать принцип суперпозиции, сущность которого заключается в следующем.
Пусть ![]() - скорость изменения плотности функции распределения числа микроорганизмов по массам (объемам) в рассматриваемой системе данный момент времени t, обусловленная «элементарным» процессом i-м, например ростом клеток, тогда скорость изменения плотности функции распределения для сложного процесса можно представить в виде суперпозиции скоростей для «элементарных» процессов (Пеньков, 1992), т.е.
- скорость изменения плотности функции распределения числа микроорганизмов по массам (объемам) в рассматриваемой системе данный момент времени t, обусловленная «элементарным» процессом i-м, например ростом клеток, тогда скорость изменения плотности функции распределения для сложного процесса можно представить в виде суперпозиции скоростей для «элементарных» процессов (Пеньков, 1992), т.е.
![]() .
.
Такой подход к математическому моделированию процессов, управляемых линейными и квазилинейными дифференциальными и интегро-дифференциальными эволюционными уравнениями математической физики, позволяет описать сложный процесс на основе кинетических уравнений, описывающих «элементарные» процессы.
Итак, пусть f(x,t) - плотность функции распределения (дифференциальная функция распределения) числа живых (способных к росту и размножению) микроорганизмов в момент времени t по массам х в единице объема дисперсной системы; fp(x,t) - плотность функции распределения числа мертвых (не способных к росту и делению) клеток по массам х в единице объема системы в момент времени t;
Г(x,t)f(x,t) - число микроорганизмов массой х, погибших в единицу времени в единице объема системы в момент времени t естественной смертью;
![]() - число клеток массой х, погибших в единицу времени в единице объема системы в момент времени t вследствие внутривидовой борьбы. Здесь Г(x,t) и G(x,ζ,t) - заданные функции своих аргументов. Тогда с учетом изложенного выше для пространственно-однородных систем, когда клетки распределены равномерно по объему, можно получить следующие квазилинейные интегро-дифференциальные уравнения, описывающие рост, размножение и гибель микроорганизмов:
- число клеток массой х, погибших в единицу времени в единице объема системы в момент времени t вследствие внутривидовой борьбы. Здесь Г(x,t) и G(x,ζ,t) - заданные функции своих аргументов. Тогда с учетом изложенного выше для пространственно-однородных систем, когда клетки распределены равномерно по объему, можно получить следующие квазилинейные интегро-дифференциальные уравнения, описывающие рост, размножение и гибель микроорганизмов:
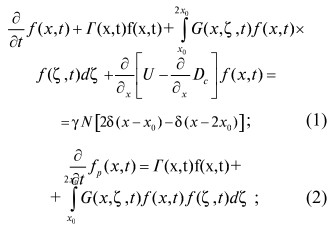
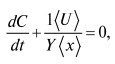 (3)
(3)
где U = U(x,C,t) - скорость роста клеток массой х в момент времени t;
Dc = Dc(x,C,t) - стохастический параметр (Dc - коэффициент диффузии в пространстве масс); γ - удельная скорость поступления в систему микроорганизмов массой х0, образовавшихся при делении клеток массой 2х0; δ(z) - дельта-функция Дирака от z; ![]() - знак среднего значения от указанной величины; Y - экономический коэффициент;
- знак среднего значения от указанной величины; Y - экономический коэффициент; ![]() и
и ![]() - соответственно масса N живых и Np мертвых клеток в единице объема системы в момент времени t.
- соответственно масса N живых и Np мертвых клеток в единице объема системы в момент времени t.
Система уравнений (1)-(3) незамкнута. Для получения дополнительной информации умножим уравнения (l) и (2) на х и проинтегрируем по х от х=х0 до х=2х0. В результате получим уравнения для определения M(t) и Мp(t):
![]() (4)
(4)
![]() (5)
(5)
Уравнения (l)-(5) следует еще дополнить законом сохранения вещества:
M(t) + Mp(t) + YC(t) = M0+YC0, Mp(0) = 0, (6)
где С0 и М0 -значения M(t) и C(t) при t = 0. Для решения уравнений (1)-(6) необходимо еще задать начальные и граничные условия:
Начальные условия:
f(x,0)= f0(x) fр(х,0)=0, М(0) = М0,
Мр(0) = 0, С(0) = С0,
где f0 (x), М0 и С0 - заданные величины
Граничные условия:
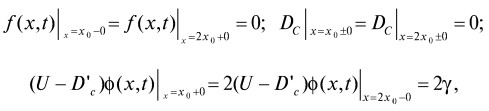
3. где ![]() предполагается, что U > D´c для всех х ∈ [х0,2х0].
предполагается, что U > D´c для всех х ∈ [х0,2х0].
4. Уравнение (1) заменой искомой функции f(x,t) на F(x,t) по формуле
f(x,t) = [θ(х -х0) -θ (х - 2x0)]F(x,t), (7)
5. с учетом того, что θ (z)=0 при z<0 и θ (z) = 1 при z>0,
dθ /dz=δ(z), zδ(z) = 0, f(x,t) δ (x - z) = f(z,t) δ(x - z),
где δ(z) - дельта-функция Дирака, преобразуется в уравнение
![]() (8)
(8)
решение которого должно удовлетворять начальному F(x,0) = F0(x) и граничному условию
![]() (9)
(9)
Аналитическое решение полученной системы уравнений представляет значительные трудности. Очевидно, что оно упрощается, если имеют место равенства:
Г(х,t)=Г0, G(x,ζ,t)=G0, U(x,C,t)=x ψ(C),
где ψ(0)=0, Г0 и G0 - постоянные величины, а ψ (С) - заданная функция своего аргумента. В этом варианте решение системы уравнений (2)-(6) можно представить в следующем виде:
M(C)=M0+Y(C0 -C) -Mp(C), C≥C*: (10)
![]() (11)
(11)
6. где ![]() Из условия М(С) = 0 при С = С* следует уравнение для определения величины С = С*:
Из условия М(С) = 0 при С = С* следует уравнение для определения величины С = С*:
M0+Y(C0-C*) = Mp(C*) (12)
Зная зависимость функции М от С, согласно уравнению (3), можно найти C(t):
![]() (13)
(13)
Выражения (10)-(13) при G0 = 0 были получены ранее (Пищиков, Пеньков, 2005).
Анализ. Из уравнения
![]()
следует, что при ψ(С) > Г0 + αМ функция M(t) возрастает от М = M0 до М=Мmах, а при ψ(С)<Г0 +αМ - убывает до М = 0. Следовательно, при ψ (Сλ) = Г0+ αМ(Сλ) функция M(t) достигает своего максимального значения. В свою очередь функция М(t) возрастает от нуля до Мp=М0 + Y(C0 - С*), причем ее график имеет точку перегиба при С=Сλ.
Из уравнения (12) следует, что при гибели клеток существует пороговая минимальная концентрация субстрата, ниже которой клеточный прирост массы живых клеток прекращается.
Однако следует заметить, что для определения величины C=C* согласно уравнениям (11) и (12), необходимо знать величину ![]() , которую, в свою очередь можно найти только в случае, если известна функция
, которую, в свою очередь можно найти только в случае, если известна функция ![]() . Очевидно, что уравнение для определения искомой функции P(x,t) можно получить, используя уравнения для функций F(x,t) и N(t):
. Очевидно, что уравнение для определения искомой функции P(x,t) можно получить, используя уравнения для функций F(x,t) и N(t):
![]() (14)
(14)
![]() (15)
(15)
Действительно, так как F(x,t) = P(x,t)N(t), то, исключая величину N(t) из уравнения (14), получим уравнения для искомых функций P(x,t) и ![]() :
:
![]() (16)
(16)
![]() (17)
(17)
Уравнение (14) справедливо и в том случае, когда ![]() , где φ(х) - заданная функция.
, где φ(х) - заданная функция.
Для решения уравнения (16) используем квазистационарное приближение, положив ![]() В этом приближении
В этом приближении
![]() (18)
(18)
Уравнение (16) значительно упрощается, когда φ(х)=х. В этом варианте в квазистационарном приближении, если левую и правую части этого уравнения умножить на х и проинтегрировать по х от х=х0 до х=2х0, используя соответствующие граничные условия, то при Dc(x, t) = U(x, t)b(x) получим достаточно компактное уравнение
![]() (19)
(19)
где ![]()
![]()
Перейдем к безразмерной переменной х= х0z. В этом случае при
![]() (20)
(20)
уравнение (19) преобразуется к виду
![]() (21)
(21)
Решение которого, удовлетворяющее граничным условиям:
![]()
![]()
![]() (22)
(22)
представимо в виде
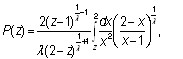 0<λ <1; (23)
0<λ <1; (23)
![]() λ=0. (24)
λ=0. (24)
Так, ![]() =2x0 ln2 при λ=0, а при λ>0 справедливо неравенство
=2x0 ln2 при λ=0, а при λ>0 справедливо неравенство
![]() (25)
(25)
Далее, так как
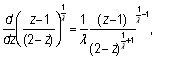
то легко показать, что функция P(z) нормирована на единицу.
В заключении следует отметить, что предложенные уравнения для описания роста, размножения и гибели микроорганизмов в закрытых пространственно-неоднородных дисперсных системах легко обобщаются на открытые системы с истоком и внешним источником микроорганизмов (см., например, Пеньков, Пищиков, 1999).
Список литературы
-
Варфоломеев С.Д., Гуревич К.Г. Биокинетика. Практический курс. М.: ФАИР-ПРЕСС, 1999. - 720 с.
-
Бейли Дж., Оллис Д. Основы биохимической технологии. Ч. I, 1989. - С. 464-457.
-
Пеньков Н.В. Коагуляционные процессы в дисперсных системах: Дис. ... докт. физ.-мат. наук. М.: НИИФХИ им. Карпова, 1992. - 342 с.
-
Пищиков Г.Б., Пеньков H.В. К кинетике роста, размножения и гибели микроорганизмов // Хранение и переработка сельхозсырья. - 2005. - №5. - С. 19-21.
-
Пеньков Н.В., Пищиков Г.Б. Кинетическая модель процесса роста, размножения и гибели дрожжевых клеток // Хранение и переработка сельхозсырья. - 1999. - №7. - С. 61-63.
Библиографическая ссылка
Пеньков Н.В. К КИНЕТИКЕ ПРОЦЕССОВ РОСТА, РАЗМНОЖЕНИЯ И ГИБЕЛИ МИКРООРГАНИЗМОВ // Успехи современного естествознания. 2011. № 1. С. 58-64;URL: https://natural-sciences.ru/ru/article/view?id=15630 (дата обращения: 20.07.2026).