



Выявление видового состава грибов-макромицетов любого региона неизбежно сталкивается с проблемой полноты изучения [6]. Пожалуй, одним из наиболее значимых факторов, препятствующим выявлению видового состава грибов, является сезонная ритмика появления их плодовых тел.
Сезонная ритмика олицетворяет одно из наиболее ярких проявлений диалектического единства организма и среды в их исторической динамике [1]. Старейшим, и до настоящего времени, основным методом накопления информации о сезонной жизни организмов остаются фенологические наблюдения – визуальная регистрация сроков появления морфологических новообразований, закономерно следующих друг за другом в процессе сезонного развития организмов. Однако фенология занимается не только наблюдениями: перед ней, кроме того, стоит задача выявления закономерностей в ходе периодического развития и определения зависимости его от условий окружающей среды, в частности определения влияния почвы и климата на периодические явления жизни растений. Фенология рассматривает местообитание как совокупность внешних влияний на организм, оцениваемых с помощью организмов–индикаторов (чаще всего – растений). Поэтому она является, по существу, инструментом экологических исследований, по крайней мере в рамках феноэкологии (Х.Л. Вернек (цит. по: [9]).
Фенология растений занимается наблюдением за заметными, легко распознаваемыми явлениями (фазами) развития и с их помощью устанавливает годовой ход развития растений. По отношению к грибам-макромицетам фенологические подходы до сих пор недостаточно проработаны. Это, в частности, связано с тем, что у этих организмов относительно легко можно учесть только одну фазу – образования плодовых тел.
Для фактического подкрепления имеющихся данных по фенологии грибов, следует, видимо, привлекать материалы по фенологии растений, с которыми эти грибы консортивно связаны. Использование этих данных становится возможным при применении принципа синхронности сезонного развития, основанного на относительной неизменности их экологии. Этот принцип означает, что на каждом этапе сезонных изменений растение нуждается в определенных экологических условиях, и, прежде всего, температуре, влажности воздуха и почвы, освещенности [5], независимо от географического положения места произрастания. Следствием этой неизменности и является синхронность развития растений, т.е. одновременность прохождения фенологических фаз.
В определенной мере данная синхронность выражена и в отношениях фенофаз растений и грибов.
Заметим, что одним из важнейших факторов, отличающих фенологию грибов от фенологии растений, является цикличность их фенофаз, т.е. в течении года некоторые виды способны образовать плодовые тела неоднократно.
Физиологическая цикличность плодоношения макромицетов в «чистом» виде может проявляться только в годы, приближающиеся по климатическим показателям к среднемноголетним. На ритм образования плодовых тел отдельных видов грибов значительное влияние оказывают погодные условия конкретного года, давая толчок к массовому плодоношению либо более ксерофильных, либо, наоборот, гигрофильных видов. Таким образом, вся группировка макромицетов в результате взаимозамещаемости отдельных видов остается стабильной и рассчитанной на максимальное обеспечение нормальной жизнедеятельности и продуктивной способности мицелия [3].
По данным Л.Г. Буровой [3], период массового плодоношения грибов-симбиотрофов составляет в среднем два года, ксилотрофов – три года. При этом ксилотрофы (и наряду с ними – гумусовые сапротрофы) отличаются наибольшей стабильностью видового состава, численности и массы; напочвенные сапротрофы адаптированы к сукцессионной этапности преобразования мертвого органического вещества и более всего подвержены погодным изменениям. Таким образом, в плодоношении макромицетов наблюдается цикличность, обусловленная, по-видимому, физиологическими особенностями состояния мицелия.
Мы поставили перед собой цель изучить феноэкологию грибов-макромицетов Южного Приуралья, так как этот регион расположен преимущественно в пределах степной зоны и его природные условия довольно экстремальны для грибов. Анализ особенностей фенологии грибов позволит лучше понять механизмы адаптации региональной микобиоты к специфическим условиям лесостепных и степных ландшафтов.
Материалы и обсуждение. Нами были проведено изучение фенологии грибов-макромицетов в разных районах Оренбургской области в период с 1993 по 2012 гг. Учет плодовых тел производился методом маршрутного учета и методом пробных площадей. На маршруте производилось описание биотопов, растительности и субстратов, на которых обитали грибы. Идентификация собранных образцов производилась с пользованием русскоязычной и зарубежной определительной литературы [2, 11-13].
В результате исследования была составлена таблица сроков плодоношения основных видов грибов региона (табл. 1).
Наши наблюдения показывают, что сезонная ритмика образования плодовых тел грибов в пределах Оренбургской области в целом соответствует данным по фенологии грибов степной зоны, которые были опубликованы Б.П. Васильковым [4]. Он утверждал, что в степной зоне выделяются два сезона образования плодовых тел грибов–макромицетов: весна и осень; в некоторые годы выделяется только один сезон плодоношения – осенний.
Исходя из наблюдений за сроками появления плодовых тел базидиальных грибов, мы выделяем пять феноритмотипов грибов–макромицетов:
- грибы образующие плодовые тела в конце весны – начале лета;
- грибы образующие плодовые тела летом;
- грибы образующие плодовые тела в конце лета и осенью;
- грибы образующие плодовые тела осенью;
- грибы образующие плодовые тела эпизодически в течение всего года;
- грибы каждой из выделенных нами фенологических групп имеют некоторые экологические особенности, общие для группы;
- грибы, образующие плодовые тела в конце весны – начале лета.
К этой группе нами отнесено 12 видов грибов: Calocera palmata, Datronia mollis, Lentinus cyathiformis, Lentinus lepideus, Lenzites betulina, Phellinus alni, Pleurotus calyptratus, Polyporus arcularius, Polyporus brumalis, Polyporus ciliatus. Грибы этой фенологической группы весьма разнородны по морфологии и систематической принадлежности. Среди этих грибов преобладают агарикоидные базидиомицеты, плодовые тела которых существуют в течение нескольких дней, а потом загнивают. Это представители родов Lentinus, Pleurotus, Polyporus. Один из наиболее типичных представителей этой группы является зимний трутовик – Polyporus brumalis.
Таблица 1
Сроки образования плодовых тел ряда грибов–макромицетов в Южном Приуралье
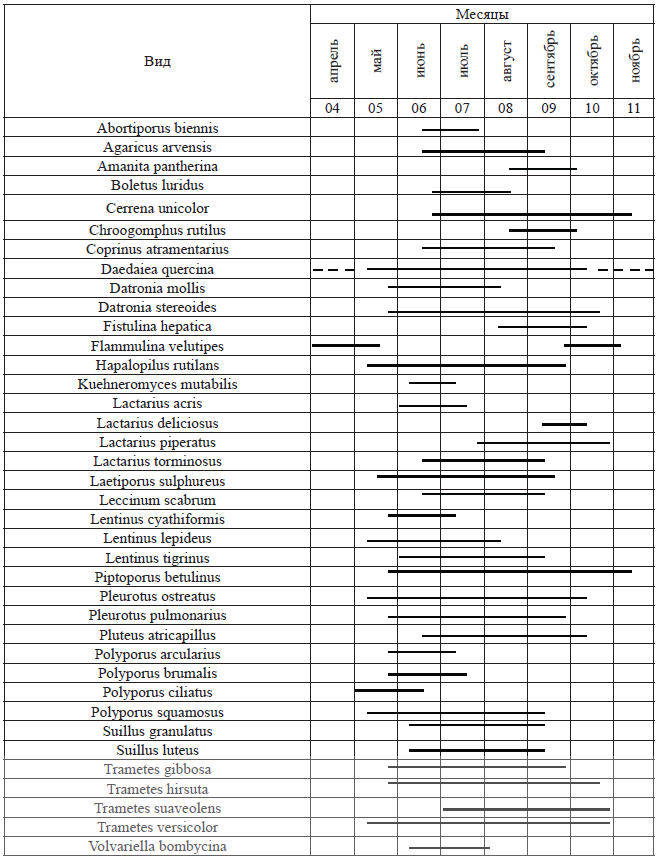
Исходя из времени образования плодовых тел можно предположить, что ключевым экологическим фактором для формирования плодовых тел данных грибов является повышенная влажность, а влияние температурного режима в данном случае менее значимо.
Грибы, образующие плодовые тела летом
К этой группе относятся 26 видов грибов – макромицетов. В частности, к этой группе относятся Abortiporus biennis, Boletus luridus, Climacodon septentrionalis, Coprinus cinereus, Daedaleopsis septentrionalis, Daedaleopsis tricolor, Hypholoma Kandoliana, Kuehneromyces mutabilis, Oudemansiella radicata, Oxyporus obducens, Pleurotus cornucopiae, Polyporus tuberaster, Russula vesca, Spongipellis spumeus, Volvariella bombycina. В эту группу входят преимущественно агарикоидные грибы, плодовые тела которых существуют несколько дней. Наиболее типичными представителями этой фенологической группы являются: Abortiporus biennis, Hypholoma Kondoliana, Kuehneromyces mutabilis, Oudenmansiella radicata, виды рода Russula. Образование плодовых тел грибов этой группы зависит в первую очередь от температурного режима. При этом сохраняется и некоторая зависимость от условий увлажнения. Исходя из резкоконтинентального климата Южного Приуралья с малым количеством летних осадков, грибы этой группы не многочисленны, и встречаются в основном в условиях повышенного увлажнения (в поймах рек, на заболоченных участках, в теневых широколиственных лесах).
Грибы, образующие плодовые тела в конце лета и осенью
Эта группа включает 64 вида грибов. К ней относятся виды родов Agaricus, Amanita, Amanitopsis, Coprinus, Hypholoma, Lactarius, Macrolepiota; а также Armillaria mellea, Chroogomphus rutilus, Clitocybe cerrusata, Clitocybe gibba, Fistulina hepatica, Hygrophoropsis aurantiaca, Leccinum scabrum, Lentinus tigrinus, Paxillus involutus, Phaeolus schweinitzii, Pleurotus dryinus, Polyporus badius, Russula foetens, Russula grisea, Scleroderma citrinum, Suillus granulatus, Trametes suaveolens. Это наиболее крупная фенологическая группа макромицетов Оренбургской области, и она очень разнородна по экологическим характеристикам входящих в нее видов. Среди них есть термофильные, ксеротолерантные виды, образующие плодовые тела в конце июня – августе, и мезофильные виды, развитие которых зависит от условий увлажнения. Большая часть видов образует плодовые тела в интервале от конца августа до начала октября, когда они собираются населением. В частности, первые маслята появляются в середине июля, однако массовое появление плодовых тел этого гриба приходится на первую неделю сентября. Похожая ситуация наблюдается и у представителей рода Xerocomus. Некоторые виды преимущественно развиваются в середине лета. Плодовые тела трутовика древолюбивого (Inocutis dryophila) появляются на живых деревьях дуба 15–20 июля и достаточно быстро разрушаются насекомыми.
Грибы, образующие плодовые тела осенью
К этой фенологической группе мы относим три вида грибов: Hebeloma crustuliniforme, Lactarius deliciosus, Lepista nuda. Можно предположить, что образование плодовых тел этих видов преимущественно зависит от условий увлажнения.
Грибы, образующие плодовые тела в течение всего года
К данной группе относятся 52 вида грибов – макромицетов Оренбургской области: Bjerkandera adusta, Daedaleopsis confragosa, Daedalea quercina, Datronia stereoides, Dichomitus squalens, Diplomitoporus flavescens, Fomes fomentarius, Fomitopsis pinicola, Ganoderma lipsiense, Gloeoporus dichrous, Gloeophyllum sepiarium, Hapalopilus rutilans, Hymenochaete tabacina, Inocutis rheades, Inocutis radiatus, Irpex lacteus, Laetiporus sulphureus, Oxyporus corticola, Phellinus igniarius, Phellinus linteus, Phellinus rhamni, Pholiota aurivella, Phellinus tremulae, виды рода Pleurotus, Polyporus squamosus, Piptoporus betulinus, Steccherinum nitidum, Steccherinum ochraceum, Schizophyllum commune, виды родов Stereum, Trametes, Trichaptum. Это вторая по количеству видов фенологическая группа. К ней относится большинство афиллофороидных грибов, особенно имеющих многолетние плодовые тела. В отношении этих видов вообще трудно определить даты начала и окончания процесса формирования плодовых тел.
Один вид – Flammulina velutires (зимний гриб) мы не можем отнести ни к одному из выделенных нами феноритмотипов. Плодовые тела этого гриба образуются только весной и осенью. Первые находки этого вида датируются 7 марта, а последние – 30 октября. Летом плодовые тела этого гриба не отмечались.
Для более полного изучения фенологии ксилотрофных грибов нами была заложена пробная площадка в пойме реки Урал в шести километрах выше по течению от г. Оренбурга. Тип леса на пробной площадке – дубрава ландышевая, Д8Ос2Тn1. Сомкнутость крон – 50–80 %. Травянистый ярус разрежен, представлен ландышем майским, ежевикой сизой, гравилатом городским, будрой плющевидной. Подрост отсутствует, в подлеске встречается жимолость татарская.
В ходе работ на пробной площадке проводилось изучение видового состава ксилотрофных базидиомицетов с периодичностью один раз в десять дней с мая по октябрь. При каждом обследовании учитывалось количество плодовых тел каждого вида гриба, их состояние и площадь гименофора. Площадь гименофора учитывалась с помощью палетки с ценой ячейки 0,25 см2. Параллельно с учетом плодовых тел велся ежедневный учет климатических показателей: влажности воздуха, минимальной и максимальной суточной температуры, среднесуточной температуры.
В течение года на пробной площадке было отмечено 32 вида грибов. В анализ были включены 20 видов, которые, судя по всему, являются типичными обитателями данного биотопа, а не случайными видами.
Анализ встречаемости грибов–макромицетов по датам наблюдений показал, что видовой состав грибного сообщества заметно варьирует в течение года (судя по наличию плодовых тел). Максимальное число видов было отмечено в конце июня и в течение всего августа (рисунок).
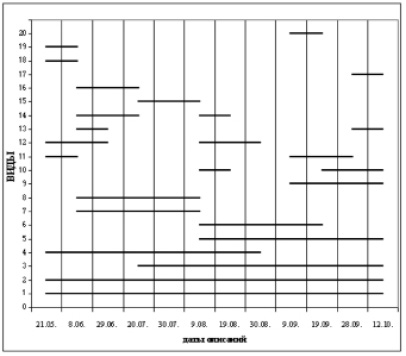
Сроки образования плодовых тел грибов на пробной площадке. Виды: 1 – Fomes fomentarius; 2 – Phellinus linteus; 3 – Irpex lacteus; 4 – Oxyporus corticola; 5 – Daedalea quercina; 6 – Fistulina hepatica; 7 – Steccherinum ochraceum; 8 – Bjerkandera adusta; 9 – Stereum hirsutum; 10 – Trametes ochracea; 11 – Trametes versicolor; 12 – Schizophyllum commune; 13 – Tremella ssp.; 14 – Pleurotus pulmonarius; 15 – Trametes pubescens; 16 – Inocutis dryophila; 17 – Exidia glandulosa; 18 – Cerrena unicolor; 19 – Laetiporus sulphureus; 20 – Hypholoma fasciculare
Одной из задач наших исследований было выяснение влияния внешних (климатических) факторов на фенологию грибов–макромицетов. В этом плане оценка влияния факторов среды на грибные организмы только по наличию или отсутствию плодовых тел не является приемлемой, поэтому в качестве маркера мы использовали площадь гименофора грибов, поскольку образование спор является важнейшей функцией плодового тела [7].
Для оценки влияния отдельных факторов на появление и развитие плодовых тел, проводился расчет коэффициентов корреляции между площадью гименофора и каждым из климатических показателей для девяти видов грибов, отмеченных на площадке. Корреляционный анализ показал, что для разных видов грибов – макромицетов ключевыми являются разные факторы (табл. 2).
Таблица 2
Ключевые факторы, определяющие формирование и развитие плодовых тел некоторых грибов пробной площадки
|
Вид |
Ключевые факторы |
Коэффициент корреляции (p<0,05) |
|
Inocutis dryophila |
Тmin |
0,89 |
|
Тср |
0,87 |
|
|
Iϕ/τ |
0,75 |
|
|
Fistulina hepatica |
Tср |
0,63 |
|
Тmin |
0,64 |
|
|
Oxyporus corticola |
ϕ |
–0,69 |
|
Bjerkandera adusta |
Тmin |
0,69 |
|
Trametes ochracea |
Iϕ/τ |
0,82 |
|
Tmax |
–0,62 |
|
|
Irpex lacteus |
Tср |
–0,66 |
|
Тmin |
–0,71 |
|
|
Steccherinum ochraceum |
Тmin |
0,77 |
|
Tremella mesenterica |
Тmin |
0,66 |
|
Stereum subtomentosum |
Iϕ/τ |
0,70 |
Примечание. T – температура (минимальная, максимальная, средняя), ϕ – относительная влажность воздуха; Iϕ/τ – гидротермический коэффициент.
Так, для Bjerkandera adusta, Irpex lacteus, Steccherinum ochraceum, Tremella mesenterica ключевым фактором является минимальная суточная температура. При этом для Irpex lacteus характерна отрицательная корреляция. Для других видов, в частности, Trametes ochracea и Stereum hirsutum, более значимым является гидротермический показатель. Образование плодовых тел Inocutis dryophila зависит от нескольких факторов, важнейшим из которых является температурный показатель.
Заключение
Отличия в фенологии разных видов грибов является одним из важнейших факторов, затрудняющих описание грибных сообществ, поскольку в результате появления плодовых тел разных видов в разное время наблюдаются как бы постоянные флуктуации их видового состава. При проведении исследований в разное время года сообщество, так сказать, «поворачивается» к нам то одной, то другой своей «гранью», скрывая при этом свое истинное лицо, не давая возможности достаточно полно оценить его видовой состав. Вследствие этого возникает вопрос: насколько оправдан анализ сообществ, видовой состав которых досконально не выявлен? В этом отношении следует опереться на опыт решения аналогичной проблемы, имеющийся в фитоценологии.
В любом случае, какие бы проблемы не возникали у исследователей в связи с фенологией грибов, для лесных сообществ, микоценозов и для самих грибов определенная ритмика образования плодовых тел является очень важным адаптивным механизмом. А. Марчелло [10] писал по этому поводу: «Способность живых организмов следовать ритмичному изменению внешней среды определяет потенциал выживания вида».
Отличия в фенологии отдельных видов грибов является одним из проявлений дифференциации их экологических ниш в сообществе, приводящим к снижению конкуренции между видами. Дальнейшие исследования в этом направлении поможет лучше понять механизмы дифференциации экологических ниш базидиальных грибов и выявить закономерности образования их базидиом. Кроме того, изучение фенологии грибов в широких масштабах может стать инструментом изучения трансформации микобиоты под влиянием климатических и антропогенных изменений [8].
Библиографическая ссылка
Сафонов М.А. ФЕНОЭКОЛОГИЯ БАЗИДИАЛЬНЫХ ГРИБОВ В УСЛОВИЯХ ЮЖНОГО ПРИУРАЛЬЯ // Успехи современного естествознания. 2013. № 8. С. 119-125;URL: https://natural-sciences.ru/ru/article/view?id=32743 (дата обращения: 01.07.2026).