



Если рассматривать систему корреляционных связей морфологических признаков как проявление целостности организма растений, то степень устойчивости корреляционных связей будет являться мерой реакции растений на различные стрессовые воздействия [4]. Под влиянием внешних условий происходят не только изменения значений отдельных признаков, но разнообразные преобразования системы взаимосвязей между этими признаками. Н.С. Ростова [5] по общей и согласованной изменчивости выделила 4 группы признаков: высоко детерминированные и сильно изменчивые (эколого-биологические); высоко детерминированные и слабо изменчивые (биологические); слабо детерминированные и слабо изменчивые (генотипические); слабо детерминированные и сильно изменчивые (экологические). На основании многочисленных исследований по изменению корреляций морфологических признаков у разных видов культурных и дикорастущих видов растений Ростова [5] сформулировала общие закономерности. Ухудшение условий среды, в том числе уменьшение площади питания, приводит к увеличению общей и согласованной изменчивости эколого-биологических признаков. В более благоприятных условиях снижается вариабельность и сила связей в этой группе признаков. У биологических признаков при ухудшении условий детерминированность увеличивается, вариабельность может увеличиваться или уменьшаться. У таксономических (генотипических) признаков низкий уровень общей и согласованной изменчивости остаётся стабильным в разных условиях среды. У экологических признаков при ухудшении условий детерминированность снижается, вариабельность может уменьшаться или увеличиваться.
Erysimum cheiranthoides L. – желтушник левкойный – перспективное лекарственное растение кардиотонического действия [1]. Создание сырьевой базы для получения препаратов кардиотонического действия значительно снизит зависимость отечественного производителя от импортных поставок. Ранее нами было изучено влияние плотности посева на показатели морфологических признаков E. cheiranthoides [2, 3]. В результате было установлено, что изучаемые морфологические признаки по степени влияния плотности посева разделились на три группы. Признаки первой группы: длина, ширина листа, длина главного побега не изменялись под влиянием плотности посева. Во вторую группу вошёл один признак длина префлоральной части побега, который увеличивал свои показатели по мере увеличения плотности посева. В третью группу вошли большинство признаков: длина флоральной части побега, число листьев, число и длина боковых побегов, масса побегов, число, масса цветков, масса листьев, масса корней, масса особи. Все признаки третьей группы значительно уменьшали свои показатели по мере увеличения плотности посева. Доля влияния плотности посева на признаки первой и второй групп составляла всего 2,0 %, а доля влияния плотности посева на признаки третьей группы значительно выше и составляла 36,2 %.
Цель настоящего исследования – определить влияние плотности посева на изменчивость силы и структуры корреляционных связей всех изучаемых морфологических признаков.
Материалы и методы исследования
Опытные посевы проводили семенами пятой репродукции весной 2009 г. Семена высевали рядовым способом, с междурядьями 70 см на ровных участках площадью 60 погонных метров для каждого варианта опыта. Семена высевали на одинаковую глубину, точно по счёту на каждый погонный метр. Испытывали 5 вариантов: 1 – 3000 шт. (масса семян 0,66 г) очень высокой; 2 – 1000 шт. (0,22 г) высокой; 3 – 500 шт. (0,11 г) средней; 4 – 200 шт. (0,044 г) низкой плотности посева и 5 – 50 шт. (0,011 г) разреженный посев. Сорняки в посевах уничтожали полностью в течение всего сезона. В каждом варианте посева отбирали по 50 особей в период цветения. Для анализа были выбраны статические признаки [4]. В число статических признаков вошли следующие признаки: длина префлоральной части побега, см (А), длина флоральной части побега, см (Б); длина главного побега, см (В); число листьев на особь, шт. (Г); длина наиболее развитого листа, см (Д); ширина листа, см (Е); число боковых побегов (паракладиев), шт. (И); длина паракладиев, см (К); число цветков на особь, шт. (Л); масса листьев, г (М); масса цветков, г (Н); масса побегов, г (О); масса корней, г (П); масса особи, г (Р).
При проведении корреляционного анализа использован подход, разработанный Н. С. Ростовой [5]. Для каждой выборки вычисляли коэффициенты корреляции между всеми признаками. На основании каждой корреляционной матрицы по методу П. В. Терентьева [6] был построен корреляционный цилиндр с сечением r = 0,7. Программа построения корреляционных колец была любезно предоставлена П. А. Лезиным (ЗИН РАН). Для оценки среднего уровня связей использовали коэффициент детерминации (квадрат коэффициента корреляции), усреднённый по всей матрице (R2m) и по отдельным признакам (R2ch). Определение сходства полных матриц или частей матриц (столбцов, соответствующих связям отдельных признаков) производили с помощью метода главных компонент по z-преобразованным матрицам. При этом первая главная компонента интерпретировалась как фактор сходства матриц, вторая компонента отражала различия между матрицами [5].
Результаты исследования и их обсуждение
По общей и согласованной изменчивости признаки E. cheiranthoides подразделяются на три группы (табл. 1). К эколого-биологическим индикаторам, показатели которых высоко детерминированы (R2ch=0,327-0,649) и сильно изменчивы (CV=59,4-201,9 %), относятся большинство продуктивных признаков: число и длина паракладиев (И, К), число листьев (Г), число цветков на особи (Л), масса листьев, цветков, побегов, корней и особи (М, Н, О, П, Р). При увеличении плотности посева (от 4 к 2 варианту) происходит сначала увеличение вариабельности и степени детерминированности большинства этих признаков (максимум наблюдается в 3-м варианте), а затем уменьшение их общей и согласованной изменчивости (вариант 2). В разреженных посевах (вариант 5) и посевах очень высокой плотности (вариант 1) подобной закономерности не наблюдается. По всей видимости, связано это с тем, что разреженные посевы – оптимальные условия для роста и развития особей E. cheiranthoides, а посевы очень высокой плотности – наиболее дискомфортные [3]. Большинство признаков эколого-биологической группы чрезвычайно вариабельны как в посевах одной плотности, так и в посевах разной плотности [1, 3]. Связано это с тем, что в посевах одной плотности встречаются особи, у которых некоторые значения признаков этой группы равны нулю. В разреженных посевах доминируют особи с максимальным числом боковых побегов, но совершенно неразвитой префлоральной частью побега. При увеличении плотности посева значительно увеличивается вариабельность большинства признаков: число, длина боковых побегов, число листьев, цветков, масса органов и особи (табл. 1). При варьировании плотности посева под действием естественного отбора появляется высокая пластичность признаков побега и листьев. Если количественные характеристики признака имеют приспособительное значение, изменчивость его становится объектом отбора и отбор диктует размах колебаний признака.
Структура связей признаков эколого-биологической группы достаточно стабильна при изменении плотности посева (FD1=72,5-89,0 %). Как отмечает Н.С. Ростова [5], эколого-биологические признаки являются индикаторами системной адаптивной изменчивости организмов в неоднородной среде, позже других заканчивают формирование в онтогенезе и образуют ядра корреляционных плеяд. Полученные нами данные согласуются с результатами этого автора. Как было показано нами ранее [3], изменения плотности посева больше всего (36 %) влияют на показатели эколого-биологических признаков.
Общая и согласованная изменчивость признаков растений E. cheiranthoides в посевах разной плотности
|
Признаки |
Плотность посева |
FD1 |
|||||||||
|
1 |
2 |
3 |
4 |
5 |
|||||||
|
CV |
R2ch |
CV |
R2ch |
CV |
R2ch |
CV |
R2ch |
CV |
R2ch |
||
|
Длина префлоральной части побега, см |
48,8 |
0,135 |
58,5 |
0,147 |
57,2 |
0,290 |
108,4 |
0,324 |
156,5 |
0,162 |
89,5 |
|
Длина флоральной части побега, см |
75,5 |
0,288 |
73,8 |
0,447 |
66,4 |
0,381 |
36,3 |
0,325 |
26,2 |
0,128 |
94,2 |
|
Длина главного побега, см |
35,1 |
0,152 |
32,3 |
0,126 |
22,9 |
0,092 |
16,8 |
0,073 |
14,4 |
0,030 |
63,6 |
|
Число листьев, шт. |
84,8 |
0,552 |
137,1 |
0,473 |
144,4 |
0,606 |
85,1 |
0,477 |
98,0 |
0,387 |
88,0 |
|
Длина листьев, см |
51,2 |
0,440 |
44,7 |
0,367 |
46,5 |
0,322 |
25,3 |
0,113 |
27,2 |
0,087 |
71,0 |
|
Ширина листьев, см |
47,8 |
0,434 |
55,0 |
0,370 |
51,0 |
0,379 |
28,3 |
0,104 |
29,3 |
0,087 |
77,8 |
|
Число боковых побегов, шт. |
146,4 |
0,536 |
132,8 |
0,521 |
131,6 |
0,584 |
83,2 |
0,491 |
114,4 |
0,468 |
86,9 |
|
Длина паракладиев, см |
201,9 |
0,480 |
168,4 |
0,515 |
161,7 |
0,622 |
87,4 |
0,527 |
131,2 |
0,476 |
86,0 |
|
Число цветков, шт. |
103,6 |
0,581 |
111,8 |
0,388 |
173,9 |
0,479 |
101,8 |
0,318 |
99,3 |
0,434 |
76,8 |
|
Масса листьев, г |
125,6 |
0,599 |
151,7 |
0,428 |
175,1 |
0,602 |
83,4 |
0,471 |
59,4 |
0,403 |
72,5 |
|
Масса цветков, г |
101,9 |
0,462 |
124,8 |
0,327 |
147,4 |
0,517 |
106,9 |
0,339 |
160,2 |
0,438 |
75,1 |
|
Масса побегов, г |
112,9 |
0,633 |
114,6 |
0,567 |
119,5 |
0,629 |
69,4 |
0,521 |
96,5 |
0,486 |
88,3 |
|
Масса корней, г |
117,3 |
0,542 |
118,9 |
0,497 |
152,9 |
0,565 |
67,9 |
0,416 |
71,9 |
0,375 |
89,0 |
|
Масса особи, г |
106,5 |
0,614 |
118,0 |
0,574 |
134,2 |
0,649 |
70,5 |
0,546 |
97,4 |
0,477 |
87,3 |
|
Итого |
97,1 |
0,480 |
103,0 |
0,420 |
113,2 |
0,480 |
84,4 |
0,360 |
69,4 |
0,296 |
84,1 |
CV – коэффициент вариации, %, R2ch – коэффициент детерминации, FD1 – степень сходства структуры связей признаков, %.
К таксономическим признакам, слабо детерминированным (R2ch=0,030-0,152) и мало изменчивым (CV=14,4-35,1 %), относится только длина главного побега (табл. 1). Как отмечает Н.С. Ростова [5], таксономические индикаторы устойчивы к внешним воздействиям и отличаются наибольшей стабильностью и автономностью в своём развитии. Ранее нами [3] было показано, что не выявлено влияние плотности посева на показатели именно этого признака. При увеличении плотности посева наблюдается закономерное увеличение общей и согласованной изменчивости длины главного побега (табл. 1). Структура связей данного признака в посевах разной плотности нестабильна (FD1=63,6 %).
Промежуточное положение по общей и согласованной изменчивости между группами эколого-биологических и таксономических индикаторов занимают такие признаки как длина префлоральной и флоральной части побега, длина и ширина листовой пластинки. При увеличении плотности посева характер поведения этих признаков различен. При увеличении плотности посева отмечается закономерное уменьшение общей и согласованной изменчивости длины префлоральной части побега и наоборот увеличение вариабельности и детерминированности флоральной части побега, длины и ширины листовой пластинки (табл. 1). Связано это с тем, что при увеличении плотности посева наблюдается увеличение длины префлоральной части побега и уменьшение флоральной его части [3]. Разнонаправленный характер изменения показателей этих признаков отражается и на общей и согласованной изменчивости.
Длина и ширина листьев относятся к признакам, зависимым от плотности посева в значительно меньшей степени, чем эколого-биологические признаки. Однако, в условиях сильного дискомфорта для развития (1 вариант), изменчивость листовой пластинки согласуется с изменчивостью эколого-биологических признаков.
Исходя из выше сказанного, следует, что большинство изученных признаков E. cheiranthoides в период цветения растений реагирует одинаково на увеличение плотности посева. Они уменьшают свои показатели, но увеличивают уровень согласованной изменчивости и размах вариабельности.
Степень сходства структуры связей по полным корреляционным матрицам всех вариантов плотности высокая (FD1=84,1 %). Ординация матриц методом главных компонент (рис. 1) демонстрирует, что сильнее всего от остальных отличается структура связей в посевах очень высокой плотности (вариант 1) и в разреженном посеве (вариант 5). Сходство структуры связей в 1 и 5 вариантах плотности посева составляет 88 %. Наиболее сходна структура связей в загущённых посевах и посевах средней плотности (варианты 2-4) (FD1=92,4-96,5 %).
Общие тенденции изменения структуры связей выявляет рассмотрение корреляционных матриц признаков в разных вариантах плотности посева (рис. 1). В вариантах 2-4 все признаки эколого-биологической группы объединяются в единую корреляционную плеяду (Г, И, К, Л, М, Н, О, П, Р) (рис. 1). Сильные связи (r > 0,9) объединяют характеристики побега (число и общая длина паракладиев) и массы частей растения (масса листьев, масса цветков, масса побегов, масса корней и масса особи). Независимыми оказались такие признаки, как длина главного побега, а также длина флоральной и префлоральной части побега, длина и ширина листовой пластинки.
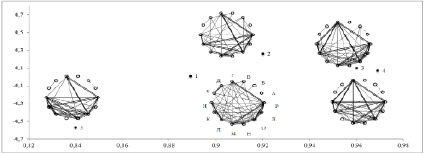
Сравнение матриц корреляций морфологических признаков E. cheiranthoides в посевах разной плотности. По оси абсцисс – фактор сходства матриц (F1); по оси ординат – фактор специфичности матриц (F2). Названия признаков даны в тексте. Условные обозначения корреляций: 0,7 < r < 0,8 – тонкая линия, 0,8 < r < 0,9 – жирная линия; r > 0,9 – двойная линия; r < 0 – пунктирная линия
В условиях очень высокой плотности посева (вариант 1) в результате очень сильной внутривидовой конкуренции происходит снижение уровня связей большинства эколого-биологических признаков плеяды (0,7 < r < 0,8): масса особи (Р), длина боковых побегов (К), масса побегов (О); число листьев (Г), масса побегов (О). Условия сильного угнетения роста и развития приводят не только к снижению уровня корреляционных связей эколого-биологических признаков, но и к перестройке в корреляционной структуре. За счет увеличения силы связей (r > 0,7) к основной плеяде примыкают признаки длина и ширина листовой пластинки.
Наиболее благоприятные для роста и развития E. cheiranthoides условия складываются в разреженных посевах (вариант 5) [3]. В условиях максимальных для развития признаков, уменьшается уровень корреляционных связей и размах изменчивости таких признаков плеяды как длина, ширина листа, число листьев, число цветков, число боковых побегов. Нарушается связь массы листьев с другими признаками. Конкуренция за свет у растений в условиях разреженного посева увеличивается во много раз за счёт конкуренции на уровне боковых побегов. Интенсивность фотосинтеза нижних листьев резко падает, прирост массы уменьшается, и связь массы листьев с другими признаками растений нарушается. Таким образом, условия разреженного посева способствуют максимальному развитию боковых побегов (число и их общая длина), что усиливает конкуренцию за свет и снижению фотосинтеза листьев нижнего яруса.
Заключение
Изучено влияние плотности посева на структуру и изменчивость корреляционных связей морфологических признаков у E. cheiranthoides. Испытывали 5 вариантов плотности посева: очень высокая (1 вариант), высокая (2 вариант), средняя (3 вариант), низкая (4 вариант) и разреженный посев (5 вариант). По общей и согласованной изменчивости выделены три группы признаков. Первую группу составляли эколого-биологические индикаторы – высоко детерминированные и сильно изменчивые признаки: длина флоральной части побега, число и общая длина паракладиев, число листьев и цветков, масса листьев, цветков, побегов, корней и особей. Структура связей признаков эколого-биологической группы достаточно стабильна при изменении плотности посева (FD1=72,5-89,0 %). Наиболее стабильными и независимыми от влияния плотности посева оказались показатели таксономического признака – длина главного побега. Промежуточное положение по общей и согласованной изменчивости занимают такие признаки как длина префлоральной и флоральной части побега, длина и ширина листовой пластинки. При увеличении плотности посева (от 4 к 2 варианту) происходит сначала увеличение вариабельности и степени детерминированности большинства эколого-биологических признаков, а затем уменьшение их общей и согласованной изменчивости (вариант 2). В этих условиях большинство признаков эколого-биологической группы объединяется в единую плеяду. В условиях сильно загущённого (вариант 1) и сильно разреженного посева (вариант 5) наблюдаются перестройки корреляционной структуры, носящие разный характер в каждом варианте. В условиях разреженного посева не обнаруживается связи между массой листьев с другими признаками. Предполагается, что в наиболее благоприятных условиях конкуренция за свет значительно возрастает, а интенсивность фотосинтеза и прирост массы в листьях нижнего яруса снижается.
Библиографическая ссылка
Годин В.Н., Гонтарь Э.М. ВЛИЯНИЕ ПЛОТНОСТИ ПОСЕВА НА КОРРЕЛЯЦИОННЫЕ СВЯЗИ МОРФОЛОГИЧЕСКИХ ПРИЗНАКОВ У ERYSIMUM CHEIRANTHOIDES L. // Успехи современного естествознания. 2014. № 7. С. 85-89;URL: https://natural-sciences.ru/ru/article/view?id=34149 (дата обращения: 01.07.2026).