



Известно, что основными объектами защитного лесоразведения являются районы с чрезвычайно жёсткими лесорастительными условиями. Недостаток влаги, часто повторяющиеся засухи, суховеи, морозные зимы, засоленность и солонцеватость почв, подверженность древостоев болезням и нападениям вредителей создают трудности выращивания долговечных насаждений. Поэтому дуб является одним из перспективных объектов для использования в защитном лесоразведении.
При отборе перспективного селекционного материала для защитного лесоразведения в сухой степи наряду с наблюдением за ростом и развитием потомства отобранного селекционного материала на устойчивость к засухе, вредителям и болезням необходимо детальное изучение физиологии и процессов органообразования. Сравнительно-морфологические наблюдения являются надежным инструментом для выявления эволюционных отношений у растений. Экспериментальное получение форм – это тот же эволюционный процесс, сжатый во времени [1, 5].
Одной из основных задач селекционеров в условиях сухой степи является отбор селекционного материала по устойчивости, долговечности и производительности путем сочетания естественного и искусственного отбора.
Морфологические и морфогенетические наблюдения позволяют более полно анализировать направления и результаты совместного действия естественного и искусственного отбора [2, 3].
Метод морфогенетического анализа базируется на представлении, что в морфогенезе растения выделяются два процесса – процесс формирования зачаточного главного побега и процесс развертывания элементов метамеров (зачатка листа, бокового конуса нарастания и узла с междоузлием) в функционирующие органы растения. В основе метода лежит деление жизненного цикла растений на этапы органогенеза, установленные на основании исследования морфологических и цитологических изменений, происходящих в клетках верхушечных меристем побегов, а также в образовательных тканях генеративных органов. При всех специфических различиях все высшие покрытосеменные проходят 12 основных этапов органогенеза, на каждом из которых происходит формирование характерных для данного этапа одноименных органов. Подробная характеристика этапов и обоснование предложенной схемы органообразовательных процессов дана Ф.М. Куперман [1, 5, 6].
Морфогенетический анализ позволяет обнаружить черты сходства и отличия в морфогенетическом цикле сравниваемых форм, выявить сбалансированность процесса морфогенеза на отдельных этапах онтогенеза, составить представление о типе растения каждой формы. Анализ сортовой популяции позволяет выявить имеющееся у растений в посеве разнообразие соотношений процессов роста и развития, выявить в пределах формы морфофизиологические группы, различающиеся по интенсивности ростовых и биометрических процессов, дает возможность получить более полную характеристику как исходного материала для отбора, так и самих отборов. Морфогенетические наблюдения дают возможность очень тонко различать состояния растений при различном сочетании факторов среды и состояние сортов и селекционных форм при одинаковых условиях выращивания, а также установить наиболее чувствительные этапы органогенеза к изменению условий.
Особую актуальность морфогенетические наблюдения имеют в специфических аридных условиях сухой степи Нижнего Поволжья [2, 4].
Методика морфологических наблюдений
Берутся пробы растений, фиксируется их структура, анализируются доступные невооруженному глазу макроморфологические признаки, а при изучении внутрипочечной фазы развития побегов используется лупа или бинокулярный микроскоп. Исследования проводятся на свежем материале. Для получения информации о сортовых особенностях вегетативной и генеративной сфер растений достаточна выборка в 25–50 экземпляров растений. Для выявления структуры сортовой популяции выборка должна быть больше. Анализ почек проводится через каждые 3–5 дней в период интенсивного развития их весной и 1–2 раза в летне-осенние месяцы. При проведении морфофизиологических исследований необходимо соблюдение некоторых принципов:
1. При отборе проб учитывать принцип репрезентативности, то есть в пробу должна входить вся совокупность растений.
2. Количество растений для анализа должно быть достаточным, чтобы при анализе каждый раз 5–10 растений их хватило до конца опыта.
3. Частота наблюдений зависит от задачи исследований и количества растений.
4. При сравнении различных объектов под бинокулярной лупой измерения необходимо проводить при одном и том же увеличении.
Объектом исследований был дуб черешчатый, представленный двумя популяциями: Чапурниковской и Арчединской. Чапурниковская байрачная популяция относится к естественным дубравам Чапурниковской балки, расположенной в южной оконечности г. Волгограда в аридных условиях на границе сухой степи и полупустыни. Арчединская популяция относится к естественным дубравам надпойменной террасы Арчединского лесхоза, занимающего территорию примерно в 200 км к северо-западу от Волгограда. На момент исследования возраст этих естественных популяций составлял 100–200 лет [4].
Результаты исследования и их обсуждение
В мае 2005 г. желуди исследуемых популяций были высеяны на одну грядку опытного питомника Нижневолжской станции ВНИАЛМИ (г. Камышин). Первый морфогенетический анализ проростков был проведен 2 июля. Всего за вегетационный период было проанализировано 39 побегов Чапурниковской популяции и 41 – Арчединской. 6 % желудей Арчединской популяции были повреждены долгоносиком и гнилью, у желудей Чапурниковской популяции – 19 %.
Работа с вегетирующими растениями при морфогенетическом анализе имеет два этапа:
1) фиксация структуры отдельных растений;
2) выявление специфики морфогенетических процессов.
Фиксация структуры – это составление карты схем растений. Анализ полученных данных карт схем растений дает возможность выявлять присущую популяции биоритмику и потенциальные возможности побегообразования, позволяет учесть популяционную вариабельность морфологических показателей растений.
Анализируемые растения условно были разбиты на 4 группы. Растения первой группы в момент анализа находились в стадии спящей почки. При рассмотрении зародыша семени под бинокулярной лупой были различимы только кроющие чешуи почки. Вторая группа растений отнесена к стадии проростка, когда кроме почечных кроющих чешуй вполне различимы неразвернувшиеся листочки. К третьей группе отнесены растения, имеющие в момент анализа хорошо развитый стебель с развернувшимися листьями и сформированной верхушечной почкой – максимально развитые растения во второй половине июня. В четвертую группу вошли растения, анализ которых проводился в середине августа. К этому времени сеянцы дали по два, а некоторые по три прироста.
Для выявления биоритмики развертывания побегов и листьев с учетом их популяционной вариабельности из карт схем растений в специальные таблицы были сведены данные по состоянию листового аппарата на главном побеге. Для этого у всех растений последовательно просматривались главные побеги и листья каждого побега по номерам метамеров. Отсчет листьев велся снизу. Отдельно отмечались развернувшиеся и неразвернувшиеся листья.
Таким образом, анализируя биоритмику развертывания листьев на главном побеге растений, отмечаем, что кроющие чешуи встречаются на главном побеге во всех группах обеих популяций до 9 и 11 метамера. Основная масса растений имеет кроющие чешуи по 7 метамер. Нераспустившиеся листья у растений II группы начинаются у растений Арчединской популяции с 7, а Чапурниковской популяции с 5 метамеров. Основная масса их у обеих популяций приходится на 10 и 11 метамер, заканчиваются на 15 и 16 метамере. Кроющие чешуи первой верхушечной почки появляются у Арчединской популяции с 12, а у Чапурниковской популяции с 9 метамера. Колебания незначительные. На ранних этапах развития растений различных популяций дифференциация проростков может выражаться лишь в различной интенсивности роста их и в различном числе развернутых на главном побеге листьев. Приходим к выводу: в стадии проростка значительных отличий между растениями двух популяций не обнаружено.
Растения II группы, имеющие развернутые листья и сформированную почку к моменту анализа, имеют кроющие чешуи до 10 метамера. Развернувшиеся листья начинаются у растений Арчединской популяции с 6, а у Чапурниковской популяции с 7 метамера, заканчиваются по 19 и 17 метамер соответственно. Основная масса развернувшихся листьев у обеих популяций приходится на 11 и 12 метамер. Процесс разворачивания листьев Арчединской популяции несколько растянут от метамера к метамеру. В среднем на одно растение Арчединской популяции приходится 7,1 листа, Чапурниковской популяции – 6,1. Однако с 13 метамера начинаются неразвернувшиеся листья, а с 15 – кроющие чешуи первой верхушечной почки. Несколько растянута встречаемость неразвернувшихся листьев – до 21 метамера – у растений Арчединской популяции, а у Чапурниковской – до 18, также кроющие чешуи верхушечной почки – по 22 и 19 соответственно.
У растений IV группы имеются незначительные отличия встречаемости развернувшихся листьев: у Арчединской популяции до 17, а у Чапурниковской популяции до 19 метамера. Несколько растянута встречаемость кроющих чешуй первой верхушечной почки у растений Чапурниковской популяции – с 13 по 25 метамер, в отличие от Арчединской – с 15 по 23. Сильно растянута встречаемость развернувшихся листьев второго элементарного побега у сеянцев Чапурниковской популяции – с 19 по 35, а у Арчединской – с 20 по 31. Но основная масса их находится на 26 и 27 метамере как у той, так и у другой популяции. Незначительна разница по остальным показателям.
Таким образом, выявление биоритмики развертывания листьев на главном побеге с учетом популяционной вариабельности по числу их позволило обнаружить некоторые отличия процессов органообразования Арчединской и Чапурниковской популяций, но незначительные.
Максимально возможная реализация морфогенеза в данных условиях выращивания проявилась у Арчединской популяции. Растения этой популяции имели небольшую среднюю высоту и количество видимых метамеров главного побега (таблица).
Минимальные и максимальные значения показателей Арчединской популяции также превосходили те же показатели Чапурниковской популяции.
Учет размеров и внешнего вида конусов нарастания пазушных почек и зачаточных междоузлий, включающий морфогенетический анализ апикальной меристемы зародышевой почки, дает представление о потенциально возможной в конкретных условиях вегетации структурной основе растений. Исследования отчетного года проводились при помощи бинокуляра Лейца, при этом удалось промерить динамику изменения размеров конусов нарастания без их зарисовки (рисунок).
Биометрические показатели сеянцев дуба черешчатого Арчединской и Чапурниковской популяций по группам развития
|
Показатели количества |
Арчединская |
Чапурниковская |
||||
|
min |
среднее |
max |
min |
среднее |
max |
|
|
I группа КЧ |
7,0 |
11,3 |
15,0 |
4,0 |
7,5 |
12,0 |
|
II группа КЧ |
6,0 |
7,6 |
9,0 |
4,0 |
13,0 |
10,0 |
|
НЛ |
4,0 |
6,3 |
9,0 |
1,0 |
4,4 |
7,0 |
|
III группа |
5,0 |
8,0 |
10,0 |
7,0 |
8,4 |
10,0 |
|
РЛ |
5,0 |
7,0 |
9,0 |
5,0 |
6,5 |
8,0 |
|
НЛ |
2,0 |
2,7 |
6,0 |
1,0 |
2,2 |
4,0 |
|
КЧ1 |
9,0 |
10,0 |
12,0 |
9,0 |
10,8 |
14,0 |
|
НЛ1 |
2,0 |
3,3 |
4,0 |
2,0 |
2,4 |
4,0 |
|
Высота, см |
12,9 |
14,1 |
15,7 |
8,1 |
11,3 |
15,3 |
|
Количество видимых метамеров |
14 |
17 |
21 |
14 |
16,6 |
18 |
|
IV группа КЧ |
8 |
8,8 |
11,0 |
8,0 |
9,0 |
11,0 |
|
РЛ |
6 |
7,0 |
8,0 |
5,0 |
7,8 |
9,0 |
|
НЛ |
1 |
|||||
|
КЧ1 |
5 |
5,8 |
7,0 |
6,0 |
6,5 |
8,0 |
|
РЛ1 |
5 |
7,2 |
9,0 |
5,0 |
7,5 |
10,0 |
|
НЛ1 |
4 |
4,8 |
7,0 |
2,0 |
3,7 |
5,0 |
|
КЧ2 |
5 |
9,2 |
13,0 |
8,0 |
13,0 |
18,0 |
|
НЛ2 |
4 |
4,8 |
6,0 |
4,0 |
5,6 |
8,0 |
|
Высота, см |
17,2 |
21,0 |
25,6 |
16,0 |
20,3 |
25,3 |
|
Количество видимых метамеров |
32,0 |
36,6 |
46,0 |
28,0 |
32,8 |
40,0 |
|
Количество метамеров верхушечной почки |
9,0 |
13,0 |
19,0 |
12,0 |
20,8 |
28,0 |
Примечание. КЧ – кроющая чешуя спящей почки; РЛ – развернувшийся лист первого элементарного побега; НЛ – неразвернувшийся лист первого элементарного побега; КЧ1 – кроющая чешуя первой верхушечной почки; РЛ1 – развернувшийся лист второго элементарного побега; НЛ1 – неразвернувшийся лист второго элементарного побега; КЧ2 – кроющая чешуя второй верхушечной почки; НЛ2 – неразвернувшийся лист третьего элементарного побега.
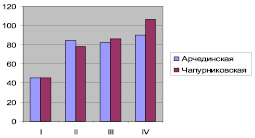
Размеры конусов нарастания растений дуба черешчатого Арчединской и Чапурниковской популяций по группам, мм
Разница между размерами конусов нарастания по группам между популяциями могла возникнуть, прежде всего, при измерении под бинокулярной лупой Лейца, имеющей грубую наводку. При подобных измерениях для увеличения точности измерений необходимо как можно большее количество наблюдений.
Проведенный ряд морфофизиологических исследований в 2005 г. на предмет выделения перспективных засухоустойчивых форм дуба не выявили отличий процессов органообразования у Арчединской и Чапурниковской популяций. Одно из положений органогенеза заключается в том, что типичный морфогенез определяется степенью оптимизации условий, необходимых для прохождения определенных этапов органогенеза. Отклонения от оптимума неизбежно приводят к появлению онтогенетических изменений морфогенеза, таких как изменение размеров метамеров, их количества, появление мутантов. В наиболее отрицательных условиях выращивания индивиды, имеющие показатели роста и развития близкие показателям морфогенеза растений, выращенных при оптимальных условиях, можно выделить как устойчивые к данному экстремуму. Таким образом, по результатам морфогенетических исследований можно сделать предварительный вывод, что Арчединская и Чапурниковская популяции достаточно устойчивы к экстремальным тяжелым почвенно-климатическим условиям сухой степи Нижнего Поволжья.
Выводы
Морфогенетический анализ, как метод морфофизиологических исследований, не выявил существенных различий в прохождении онтогенеза между различными популяциями дуба черешчатого в условиях сухой степи Нижнего Поволжья, что является результатом воздействия тяжелейших почвенно-климатических условий. Это, возможно, и приводит к нивелированию онтогенетических процессов.
Таким образом, при экстремальном ухудшении климатических характеристик региона в результате глобального потепления потомства как Чапурниковской, так и Арчединской популяций можно рекомендовать для введения в различные типы защитных лесных насаждений Нижнего Поволжья.
Библиографическая ссылка
Морозова Е.В., Иозус А.П. ВЫДЕЛЕНИЕ ЗАСУХОУСТОЙЧИВЫХ СОРТОВ ПОПУЛЯЦИЙ ДУБА ЧЕРЕШЧАТОГО ПУТЕМ МОРФОФИЗИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ В УСЛОВИЯХ СУХОЙ СТЕПИ НИЖНЕГО ПОВОЛЖЬЯ // Успехи современного естествознания. 2016. № 10. С. 90-94;URL: https://natural-sciences.ru/ru/article/view?id=36159 (дата обращения: 08.08.2026).