



В последние годы изучение углекислотного газообмена древесных пород вызывает повышенный интерес, поскольку леса, по мнению многих исследователей, имеют существенное значение в изменении климата за счет способности поглощения в процессе фотосинтеза углерода и его длительного депонирования [1].
В регуляции газового состава атмосферы значительная роль принадлежит бореальным лесам: в их растительности и верхнем слое почв сосредоточено более 22 % глобальных запасов резервуара углерода суши [2].
На территории Валдайского национального парка, который относится к одной из наиболее крупной ООПТ европейской части России, бореальные леса занимают 86 % всей площади. Особую ценность представляют высокобонитетные сосняки и ельники в возрасте 100 и более лет. Исследования ряда авторов показали, что старовозрастные леса являются хранилищем большого количества углерода, и углеродный баланс в них, как правило, положительный [3, 4].
В связи с этим изучение углекислотного газообмена ели европейской (Picea abies), одной из основных лесообразующих пород старовозрастных лесов Валдая, является актуальным, результаты могут быть использованы для количественной оценки поглощения углерода таежными лесами.
Материалы и методы исследования
Исследования СО2-газообмена проводили на экспериментальном полигоне, расположенном на территории национального парка «Валдайский» Новгородской области, – пробной площади ПП1 (размером 25 м×40 м), заложенной в старовозрастном (110–120 лет) ельнике, в течение тёплых сезонов 2013–2015 гг. Средний диаметр ели составлял 37 см, средняя высота – 31 м, бонитет – 1, формула древостоя – 9Е 1С.
Сезон наблюдений 2013 г. оказался наиболее теплым из трех сезонов – средняя температура за сезон составила 15,5 °С. Самым теплым месяцем в 2013 г. был июнь – среднесуточная и максимальная температура воздуха равнялись 18,6 и 24,2 °С.
Температурный режим 2014 г. характеризуется жарким июлем – средняя за сутки и максимальная температура воздуха – 19,5 и 25,4 °С, средняя температура за сезон – 15,2 °С.
Для более холодного сезона наблюдений в 2015 г., когда средняя температура за сезон равнялась 14,4 °С, пик показателей температуры пришелся на август – 16,3 и 21,8 °С.
Минимум температуры, в районе 6,5 °С, отмечен в зависимости от сезона в разные месяцы: в 2013 и 2014 г. – в сентябре; в 2015 г. – в мае.
По количеству осадков 2013 г. являлся наиболее дождливым годом, количество осадков в 1,5 раза превысило показатели 2014 и 2015 г. При этом максимум осадков в 2013 и 2015 г. наблюдался в июле, в 2014 г. – в июне и августе.
Облачность в наблюдаемые месяцы каждого сезона варьировала в пределах от 6 до 8 баллов. Средняя величина облачности и фотосинтетически активной радиации (ФАР) за сезон от года к году изменялась незначительно и составила 7,5 баллов и 235 мкЭ/м2.с.
Наблюдения проводили в полуденные часы в течение трех дней последней декады каждого месяца, в трёх повторностях с интервалом 10 мин. Объектами изучения углекислотного газообмена ели служили интактные побеги нижней части крон модельных деревьев.
Интенсивность нетто-фотосинтеза и светового дыхания определяли газометрическим методом, модифицированным применительно к объекту исследования (метод закрытых камер). Комплект оборудования, специально сконструированный для полевых измерений на биологическом факультете МГУ имени М.В. Ломоносова, включал:
1) портативный инфракрасный газоанализатор CO2, смонтированный на базе инфракрасного сенсора AZ 7752 с разрешающей способностью 1 ppm (AZ Instrument Corp., Тайвань), и помпу Е 134-11-120 (Hargraves Thechnologies Corp., США);
2) камеру из прозрачного пластика объемом 5100 см3 (34×10×15 см), снабженную воздуховодными полиуретановыми трубками и вентилятором;
3) крышку для создания герметизации камеры;
4) термометр Chektemp1.
Прибор и корпус камеры образовывали замкнутую систему. Измерения выполнялись в последовательности согласно методике авторов [5]. Для получения величин светового дыхания камеру затемняли с помощью чехла из светонепроницаемой ткани. Длительность экспозиции на каждом побеге составляла 30–60 с. Интенсивность фотосинтеза и дыхания хвои ели рассчитывали по уравнению Менделеева – Клапейрона [6].
Дополнительно с исследованиями углекислотного газообмена проводили наблюдения за температурой воздуха окружающей среды и температурой воздуха в камере (значения отличались между собой не более чем на 1–2 °С), определяли общую облачность. Значения фотосинтетически активной радиации рассчитывали как функцию потенциальной суммарной солнечной радиации и облачности [7].
При обработке данных использовали корреляционный и регрессионный методы анализа.
Цель исследования
Цель наших исследований состояла в оценке углекислотного газообмена старовозрастного древостоя ели европейской (Picea abies L.) на территории национального парка «Валдайский».
Результаты исследования и их обсуждение
Исследование сезонной динамики интенсивности фотосинтеза хвои ели европейской в зависимости от факторов внешней среды (температуры воздуха и освещенности) проводили как отдельно для каждого года наблюдений, так и по средним значениям за три года. Анализ данных показал наличие прямой зависимости между показателями для всех временных периодов.
Согласно нашим расчетам, в 2013 г. максимальные и минимальные значения сезонной интенсивности нетто-фотосинтеза у ели наблюдались в июне и сентябре при температуре воздуха 25 и 11 °С и составили 9,4 и 3,4 мг СО2/г сухой массы·ч соответственно.
В 2014 г. максимум интенсивности фотосинтеза – 8,4 мг СО2/г сухой массы·ч – приходился на наиболее теплый месяц – июль, когда температура воздуха составила 24 °С; сезонный минимум интенсивности, равный 3,6 мг СО2/г сухой массы·ч, отмечен в сентябре при 10 °С.
В 2015 г. максимальное значение интенсивности нетто-фотосинтеза у ели, равное 9,0 мг СО2/г сухой массы·ч, зарегистрировано в августе при 23 °С. Понижение температуры воздуха в сентябре до 14 °С привело к снижению интенсивности фотосинтеза до 4,1 мг СО2/г сухой массы·ч.
Следует отметить, что более теплая погода в последней декаде сентября в 2015 г. способствовала увеличению интенсивности фотосинтеза по сравнению с предыдущими годами.
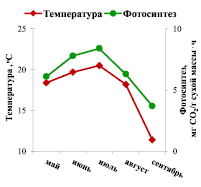
Анализ сезонной динамики интенсивности фотосинтеза хвои по средним значениям показателя за три года подтвердил результаты, полученные для каждого года: зависимость фотосинтеза ели от температуры имеет одновершинный характер – в июле отмечается пик, приуроченный к периоду высокой температуры воздуха; минимальное значение показателя зарегистрировано в сентябре (рис. 1, А).
В результате выполненного корреляционного анализа связи интенсивности фотосинтеза хвои ели и температуры воздуха также выявлена прямая линейная зависимость между показателями (коэффициент корреляции равен 0,860) [6].
Таким образом, температура воздуха является одним из основных факторов внешней среды, определяющих процесс фотосинтеза ели европейской в природных условиях.
Другим фактором, оказывающим существенное влияние на процесс фотосинтеза, является освещенность. Согласно полученным результатам, интенсивность фотосинтеза хвои ели находилась в прямой зависимости от ФАР, значения которой варьировали в диапазоне от 300 мкЭ/м2·с в сентябре до 600–640 мкЭ/м2.с в мае-июле каждого сезона.
Аналогичная связь между показателями отмечена для их среднегодовых величин (рис. 1, Б). Повышение интенсивности фотосинтеза в июле при одновременном снижении освещенности, вероятно, обусловлено превалирующим влиянием на процесс фотосинтеза температуры воздуха. Коэффициент корреляции между фотосинтетической активностью хвои и ФАР в сезонной динамике равен 0,704 [6].
Полученные нами данные согласуются с результатами ряда исследователей. Так, линейный характер зависимости скорости дневного поглощения СО2 от солнечной радиации и температуры для коренных еловых лесов Севера установлен К.С. Бобковой и В.В. Тужилкиной [8]. Согласно Г.Г. Суворовой и др., сезонный максимум фотосинтеза у ели в условиях Предбайкалья наблюдался в июле при температуре 20–25 °С, в сентябре с понижением температуры интенсивность фотосинтеза снижалась [9].
 А
А 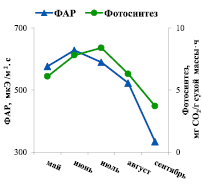 Б
Б
Рис. 1. Сезонная динамика интенсивности фотосинтеза хвои ели и факторов внешней среды: А – температуры воздуха; Б – освещенности (ФАР – фотосинтетически активная радиация) (средняя величина показателей за три года)
Вероятно, что такая зависимость между показателями обусловлена чувствительностью к действию температуры реакции карбоксилирования. Ранее нами было установлено, что диапазон оптимальных температур для проявления максимальной активности ключевых ферментов фотосинтетического углеродного метаболизма (рибулозо-1.5-бисфосфаткарбоксилазы и глицеральдегидфосфатдегидрогеназы) составляет 22–25 °С [10].
Результаты исследования интенсивности СО2-ассимиляции модельных деревьев были применены для построения многофакторной модели воздействий метеорологических условий на способность хвои ели поглощать СО2 (коэффициенты множественной корреляции и детерминации равны 0,906 и 0,820 соответственно) [6], которая послужила основой для расчета ежедневных с апреля по сентябрь каждого сезона значений интенсивности фотосинтеза в полуденные часы и, как следствие, для оценки ежедневной продуктивности фотосинтеза. При этом абсолютно сухую биомассу хвои ели определили с помощью аллометрического уравнения, в зависимости от диаметра и высоты деревьев [11], и приняли равной (с учетом категории состояния древостоя) 10,75 т на 1 га.
Фотосинтетическую продуктивность хвои за месяц рассчитывали как сумму полуденной продуктивности за все дни в каждом месяце (рис. 2).
Анализ сезонной динамики продуктивности хвои показал, что ее максимальные значения отмечены в июне 2013 г., июле 2014 г. и августе 2015 г., что вполне соотносится с сезонным ходом полуденной температуры воздуха, максимум которой приходится на обозначенные месяцы периода исследований. В сентябре для всех сезонов наблюдается уменьшение температуры воздуха и ФАР, что ведет к снижению продуктивности до минимальных значений. В целом продуктивность фотосинтеза за месяц в течение 2013–2015 гг. изменяется в пределах от 1,23 до 2,92 т СО2 /га.
Сезонную продуктивность хвои определяли как сумму продуктивности фотосинтеза за все месяцы наблюдений. Как показал анализ данных, в зависимости от метеорологических условий сезона существенных колебаний фотосинтетической продуктивности за исследуемый период не отмечается: ее показатель составляет 11,05 ± 0,26 т СО2 /га.
Таким образом, ель европейская, имеющая абсолютно сухую массу хвои 10,75 т на 1 га, синтезирует за сезон 11 т СО2 /га.
Одной из основных задач при изучении углекислотного газообмена является получение количественной характеристики взаимосвязи между фотосинтезом и дыханием, позволяющей оценить роль ели в углеродном балансе лесных экосистем. Как показали исследования, сезонный ход дыхательного газообмена хвои ели соответствует изменению интенсивности нетто-фотосинтеза: максимальные и минимальные величины дыхания, равные 3,1 и 1,6 мг СО2/г сухой массы·ч, отмечены в июле и сентябре соответственно (рис. 3). По нашим расчетам, среднее значение фотосинтеза за сезон по данным за три года, равное 6,5 мг СО2/г сухой массы·ч, в 2,7 раза превышает световое дыхание, что свидетельствует о положительном углекислотном газообмене хвои ели европейской нижней части полога старовозрастного древостоя в лесных системах Валдая.
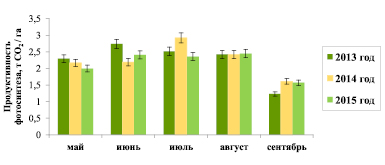
Рис. 2. Продуктивность нетто-фотосинтеза хвои ели европейской
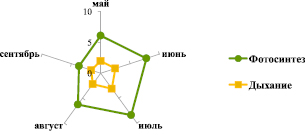
Рис. 3. Сезонные показатели фотосинтеза и дыхания хвои ели европейской (средняя величина показателей за три года, мг СО2 /г сухой массы·ч)
Заключение
Полученные результаты свидетельствуют о том, что температура воздуха и освещенность являются основными метеорологическими факторами, определяющими процесс фотосинтеза ели европейской старовозрастного древостоя таежных лесов Валдая. Линейный коэффициент корреляции между интенсивностью фотосинтеза и температурой и ФАР, равный 0,860 и 0,704 соответственно, свидетельствует о тесной связи между показателями.
Сезонный максимум нетто-фотосинтеза хвои ели европейской при температуре воздуха 23–25 °С и уменьшение поглощения СО2 из атмосферы с понижением оптимума температуры обусловлены чувствительностью реакции карбоксилирования к действию температуры.
Старовозрастные ельники Валдая имеют существенное значение, сохраняющееся с их возрастом, в углеродном балансе таежных лесов европейской части России. Подтверждением служат данные о сезонной продуктивности нетто-фотосинтеза ели европейской в наиболее благоприятных условиях освещения и температуры – в полуденные часы, а также положительный углекислотный газообмен в течение теплых сезонов 2013–2015 гг.
Авторы благодарны А.С. Маруничу (Валдайский филиал Государственного гидрологического института) за помощь в организации полевых работ и предоставление современной информации по метеостанции Валдай.
Работа выполнена в рамках темы биологического факультета МГУ имени М.В. Ломоносова № 01200117369 «Оценка состояния экосистем по данным экологического мониторинга» и при финансовой поддержке Российского научного фонда (проект № 16-17-00123).
Библиографическая ссылка
Юзбеков А.К., Иващенко А.И., Куманяев А.С. ОЦЕНКА УГЛЕКИСЛОТНОГО ГАЗООБМЕНА ЕЛИ ЕВРОПЕЙСКОЙ В ЛЕСНЫХ СООБЩЕСТВАХ ВАЛДАЯ // Успехи современного естествознания. 2017. № 6. С. 118-122;URL: https://natural-sciences.ru/ru/article/view?id=36510 (дата обращения: 30.07.2026).