



Изменения климата влияют на растительность Земли. В Финляндии имеется представление о реакции молодых лиственных деревьев на повышение температуры воздуха во взаимодействии с тропосферным озоном. Эти знания повышают шансы на разработку моделей для включения параметров, которые описывают лесную систему в изменяющихся климатических условиях [1]. Можно утверждать, что будущее сдерживание количества СО2 находится в увеличении площади, прежде всего, лесов [2, 3]. По вейвлетам универсальной конструкции изменяется динамика углерода в Европе [4, 5]. В Берлине [6] по 252 деревьям липы на кернах по расстояниям от центра к периферии были выявлены изменения приращения толщины деревьев за 50–100 лет.
Приходит понимание о моделировании взаимных связей между параметрами структуры листьев растений методом идентификации [7, 8]. Априори ясно, что именно погода влияет на ход развития и роста (онтогенеза) растений. А на многолетние растения погода влияет через ежегодный онтогенез листвы. Кванты поведения листьев, например, березы повислой, распространенной на Северном полушарии [9], четко зависят от квантов (асимметричных вейвлетов [4, 5]) поведения температуры воздуха и относительной влажности. Метеорологические условия являются сильными факторами активности биологических объектов, и для этого в статье [10] оценено влияние температуры, осадков, атмосферного давления и влажности на фенологию земноводных в Юго-Восточном Квинсленде (Австралия).
Рост растений – сложный процесс, в его основе лежат такие фундаментальные явления, как ритмичность, полярность, дифференциация, раздражимость, корреляция. Эти процессы являются общими для онтогенеза. Онтогенез – индивидуальное развитие организма от зиготы (или вегетативного зачатка) до природной смерти. Из-за фотосинтетической активности листьев растение приобретает ряд черт, которые характеризуют его рост. В процессе онтогенеза растения рост наблюдается на протяжении основных этапов его жизненного цикла [11, 12].
Формирование и отмирание листьев в цикле онтогенеза разделяются на такие этапы: распускание почек, рост и развитие листьев, расцвечивание отмирающих листьев, опадение листьев. Мы предлагаем еще два этапа онтогенеза – роста до максимума и спада до опадения.
Вегетационный период становится одним из важных экосистемных процессов, так как развитие листьев очень чувствительно к температуре воздуха. Поэтому будущее климата в наблюдениях за листьями. Метрические параметры листьев зависят от периода вегетации.
Береза повислая в Эстонии оказалась эффективной против засухи 2010 г. Деревья хорошо приспосабливаются, а значимость относительной влажности даже выше по сравнению с температурой воздуха [13]. Повышение влажности воздуха снижает температуру и накопление биомассы в молодых березках, в особенности восприимчивы листья [14].
Цель статьи: повышение точности индикации по патенту 2606189 на изобретение качества окружающей листья березы локальной среды сбоку на высоте 1,5–2,0 м со стороны преобладающих ветров по динамике средней ширины у 10 учетных листьев в различных экологических условиях произрастания около дороги с интенсивным движением автомобилей.
Материалы и методы исследования
Наибольшее время вегетации с момента распускания почек (2 мая 2014 и 1 мая 2018 г.) до максимума ширины листьев прошло 111 суток в 2014 г. (20 августа) и 110 суток в 2018 г. (18 августа). Для этапов роста (рис. 1) принимаем даты с 01.05 по 20.08 и через три часа выпишем данные «Направление ветра (румбы) на высоте 10–12 м над поверхностью земли» для метеостанции г. Йошкар-Олы. Затем клетки с «Штиль, безветрие» были исключены из массива данных, а румбы были преобразованы в азимуты.
За четыре года направление преобладающих ветров сместилось с 337,50 до 2700. Тогда можно утверждать, что направление ветров находится около северо-запада.
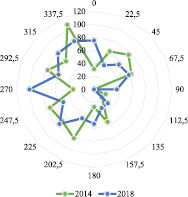
Рис. 1. Роза ветров в г. Йошкар-Оле на этапе роста в период вегетации березы повислой с 01.05 по 20.08
Данные по графикам на рис. 1 приведены в табл. 1. Здесь же даны остатки (абсолютная погрешность), как разница между фактическими и расчетными по волновым уравнениям (табл. 2). А также в табл. 1 приведены значения относительной погрешности, как отношения (в процентах) в виде деления остатков на фактические значения азимута ветров.
Колебания (вейвлет-сигналы) записываются волновой формулой [14–17] вида
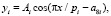
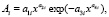
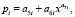 (*)
(*)
где y – показатель (зависимый фактор), i – номер составляющей модели (*), m – количество членов в модели (*), x – объясняющая переменная (влияющий фактор), a1...a8 – параметры модели (*), принимающие числовые значения в ходе структурно-параметрической идентификации в программной среде CurveExpert-1.40 (URL: http://www.curveexpert.net/), Ai – амплитуда (половина) вейвлета (ось y), pi – полупериод колебания (ось x).
Результаты исследования и их обсуждение
В табл. 2 даны параметры (*) по данным табл. 1.
Таблица 1
Направления ветров на этапе роста листьев березы повислой
|
Азимут φ, град |
Количество N румбов направлений ветра, шт. |
|||||
|
2014 г. |
Остатки |
Погрешность |
2018 г. |
Остатки |
Погрешность |
|
|
0 |
31 |
–0,210962 |
–0,68 |
75 |
0,104856 |
0,14 |
|
22,5 |
63 |
0,50304 |
0,80 |
40 |
–0,0268786 |
–0,07 |
|
45 |
75 |
–0,714944 |
–0,95 |
54 |
0,0306427 |
0,06 |
|
67,5 |
62 |
0,227208 |
0,37 |
58 |
–0,0357718 |
–0,06 |
|
90 |
0 |
0,196342 |
– |
35 |
0,0314459 |
0,09 |
|
112,5 |
19 |
–0,00369648 |
–0,02 |
0 |
–0,0262315 |
– |
|
135 |
17 |
–0,101437 |
–0,60 |
30 |
0,0266313 |
0,09 |
|
157,5 |
54 |
0,227061 |
0,42 |
31 |
0,000856083 |
0,00 |
|
180 |
27 |
0,590869 |
2,19 |
53 |
0,00342509 |
0,01 |
|
202,5 |
81 |
0,0897795 |
0,11 |
48 |
–0,0138827 |
–0,03 |
|
225 |
69 |
0,0673064 |
0,10 |
75 |
0,00819107 |
0,01 |
|
247,5 |
73 |
–0,0758387 |
–0,10 |
51 |
–0,011106 |
–0,02 |
|
270 |
32 |
–0,306591 |
–0,96 |
99 |
0,0205141 |
0,02 |
|
292,5 |
77 |
0,0334596 |
0,04 |
59 |
–0,0272314 |
–0,05 |
|
315 |
61 |
–0,0182502 |
–0,03 |
78 |
0,0145546 |
0,02 |
|
337,5 |
107 |
–0,455811 |
–0,43 |
80 |
–0,0386905 |
–0,05 |
|
Всего |
848 |
866 |
||||
Таблица 2
Параметры (1) направлений ветров по азимутам на этапе роста листьев березы
|
Номер i |
Вейвлет |
Коэф. кор. r |
|||||||
|
Амплитуда (половина) колебания |
Полупериод колебания |
Сдвиг |
|||||||
|
a1i |
a2i |
a3i |
a4i |
a5i |
a6i |
a7i |
a8i |
||
|
Этап роста вегетационного периода 2014 г. |
|||||||||
|
1 |
31,04888 |
0 |
–0,0022750 |
1 |
0 |
0 |
0 |
0 |
0,8559 |
|
2 |
4,06016e–18 |
26,00798 |
56,02962 |
0,11207 |
0 |
0 |
0 |
0 |
|
|
3 |
43,62256 |
0 |
0,00025496 |
1,51089 |
–439,5006 |
479,37559 |
0,016749 |
1,61580 |
|
|
4 |
–0,24363 |
1,10734 |
0,0094773 |
1 |
41,19066 |
0 |
0 |
1,12850 |
0,5645 |
|
5 |
8,84433e–7 |
3,96058 |
0,011097 |
1,10024 |
26,21567 |
–0,00019902 |
1,67304 |
1,62260 |
0,9919 |
|
6 |
–3,00032e–26 |
13,75107 |
0,063782 |
1,00350 |
43,87951 |
0,00014347 |
1,84964 |
0,061472 |
0,7472 |
|
7 |
4,56629 |
0 |
0,014893 |
1 |
72,84731 |
–0,013148 |
1 |
–1,08682 |
0,9478 |
|
Этап роста вегетационного периода 2018 г. |
|||||||||
|
1 |
84,77647 |
0 |
0,011844 |
1 |
0 |
0 |
0 |
0 |
0,8895 |
|
2 |
9,43003e6 |
41,69824 |
96,48688 |
0,16659 |
0 |
0 |
0 |
0 |
|
|
3 |
32,51765 |
0 |
0,0071364 |
1 |
45,13786 |
0,00018205 |
2,04081 |
4,43375 |
|
|
4 |
–4,32383e–13 |
7,19338 |
0,0060364 |
1,29657 |
27,72950 |
–0,00045090 |
1,62188 |
–4,77571 |
0,9865 |
|
5 |
–1,35019 |
0 |
–0,00039237 |
1,34949 |
23,24999 |
0,057855 |
0,99288 |
5,47970 |
0,9551 |
|
6 |
–0,36915 |
0,035157 |
0,00012797 |
1,36144 |
24,16514 |
0 |
0 |
–0,31166 |
0,4996 |
|
7 |
–1,41559 |
0,047326 |
0,0062964 |
1 |
0,23369 |
0,024887 |
1 |
5,71216 |
0,7413 |
|
8 |
4,71860 |
17,24510 |
0,073747 |
1,00392 |
22,64515 |
0,027914 |
0,98229 |
1,69285 |
0,9329 |
|
9 |
1,71127e–6 |
5,45479 |
0,56047 |
0,61560 |
722,97994 |
6,84720 |
1,18444 |
–1,45528 |
0,9157 |
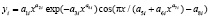
|
|
|
|
Двухчленный тренд |
Первое колебание |
|
|
|
|
Двухчленный тренд и одно колебание |
Второе колебание |
|
|
|
|
Третье колебание |
Четвертое колебание |
|
|
|
|
Пятое колебание |
Остатки после пятого колебания |
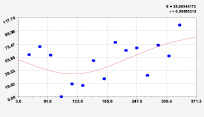
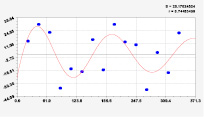
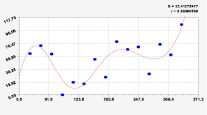
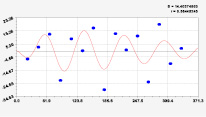
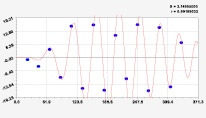
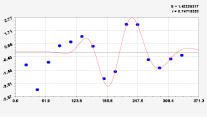
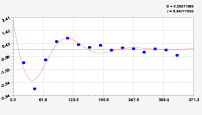
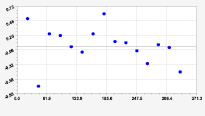
Рис. 2. Графики вейвлетов направлений ветров в 2014 г. на этапе роста листьев березы повислой
|
|
|
|
Двухчленный тренд |
Первое колебание |
|
|
|
|
Двухчленный тренд и одно колебание |
Второе колебание |
|
|
|
|
Третье колебание |
Четвертое колебание |
|
|
|
|
Пятое колебание |
Шестое колебание |
|
|
|
|
Седьмое колебание |
Остатки после седьмого колебания |
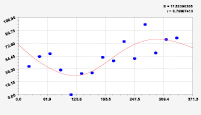
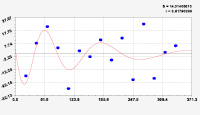
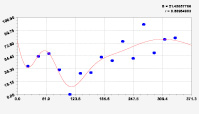
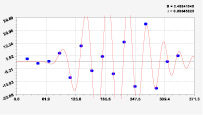
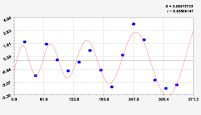
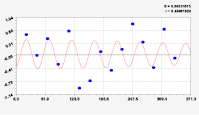
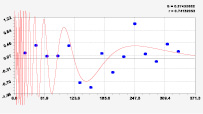
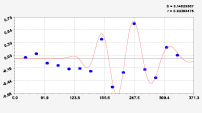
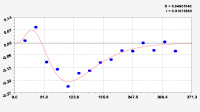
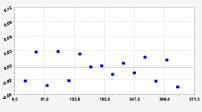
Рис. 3. Графики вейвлетов направлений ветров в 2018 г. на этапе роста листьев березы повислой
Первый член моделей с параметрами из табл. 2 является законом Лапласа (в математике), Мандельброта (в физике), Ципфа – Перла (в биологии) и Парето (в эконометрике). Но разной направленности: для 2014 г. это закон экспоненциального роста, а для 2018 г. – экспоненциальной гибели. Этот естественный закон изменяется для направлений ветров в зависимости от азимута с коэффициентами корреляции 0,5865 и 0,8000. Этот факт указывает на то, что этап роста вегетационного периода 2018 г. был лучше по направлениям ветров. Второй член тренда является биотехническим законом [14]. Все колебания имеют очень высокую адекватность. Именно это свойство колебаний указывает на то, что растения за сотни миллионов лет эволюции адаптировались к волновым изменениям метеорологических параметров.
На рис. 2 и 3 показаны графики всех составляющих общей модели (*).
Максимальная относительная погрешность по табл. 1 равна 2,19 % для 2014 и 0,14 % для 2018 г. При этом первые три члена, полученные по возможностям программной среды CurveExpert-1.40, получили коэффициент корреляции 0,8559 и 0,8859. Все пять колебаний на этапе роста 2014 г. завершились до 20 августа, когда был максимум средней ширины.
Для этапа роста в 2018 г. (рис. 3) получились семь колебаний.
Из них только четвертое колебание продолжилось после 20.08. Пятое колебание показывает, что вегетационный период начался по направлениям ветров спонтанно, а затем период колебания увеличился.
Закон стрессового возбуждения по второму члену из табл. 2 показывает, что, по-видимому, направление ветра стимулирует рост вегетационных органов растений. Этому способствовало третье колебание (пятый член) с коэффициентом корреляции 0,9919 в 2014 г. А в 2018 г. усиление колебательной адаптации роста листьев произошло из-за второго члена (биотехнический закон) и четвертого члена сильнейшей адекватности (0,9865).
Таким образом, нами доказано, что трехчасовое распределение направлений ветра по азимуту на первом этапе вегетационного периода листьев березы повислой от 1 мая до 20 августа каждого года происходит по четким закономерностям, в том числе и волновым уравнениям с переменной амплитудой, изменяющейся по биотехническому закону.
При этом период колебаний изменяется в широком диапазоне. Пока непонятно, почему первое колебание в 2014 г. происходило с отрицательным периодом 2×(–439,5) = –879 суток. Остальные волны имели периоды в 82,4; 52,4; 87,8 и 145,7 суток. Полупериод 41,2 сутки постоянный на весь этап роста листьев в 2014 г. и близок к примете в 40 суток погоды.
Для 2018 г. образовалось семь волн с полупериодом колебаний в 45,1; 27,7; 23,25, 24,17; 0,23; 22,65 и 723 суток. Из них постоянным является период в 2×24,17 = 48,3 суток. Старт учетных листьев по последнему колебанию показывает, что максимальный полупериод на 01.05.2018 у направлений ветра равен 723 суток. Этап роста листьев березы повислой в 2018 г. получил большую запутанность по сравнению с 2014 г. из-за возникновения пятого и седьмого колебаний. Такие неопределенности, по-видимому, будут нарастать на будущее.
Анализ трехчленных графиков показывает, что в 2014 г. возникли три преобладающих направления ветра, по возрастанию частоты встречаемости азимута ветров: 45; 202,5 и 337,50. А в 2018 г. образовались только два направления преобладающих ветров: 67,5 и 2700. Характерно, что из 848 направлений ветра на этапе роста листьев березы в 2014 г. не было ни одного случая по направлению азимута 900, а в 2018 г. ноль событий из 866 – по азимуту 112.50.
Заключение
Таким образом, этап роста листьев березы повислой до максимальной ширины (аналогично и длины, площади и периметра листьев) вегетационного периода становится великолепным квантом поведения растений. Этот квант с высокой адекватностью показывает, что по трехчасовым измерениям азимута ветров распределение количества направлений ветра происходит по сверхсильным закономерностям в 2014 и 2018 г. Тогда трехчасовые кванты измерений азимута ветров позволяют с очень высокой адекватностью определить распределения количества измерений направлений ветров.
В 2014 г. возникли три преобладающих направления, по росту встречаемости азимута: 45; 202,5 и 337,50. А в 2018 г. были два направления преобладающих ветров: 67,5 и 2700.
Библиографическая ссылка
Мазуркин П.М., Кудряшова А.И. ЗАКОНОМЕРНОСТИ РАСПРЕДЕЛЕНИЯ АЗИМУТА ВЕТРОВ НА ЭТАПЕ РОСТА ЛИСТЬЕВ БЕРЕЗЫ ПОВИСЛОЙ В ПЕРИОДЫ ВЕГЕТАЦИИ 2014 И 2018 ГОДОВ // Успехи современного естествознания. 2019. № 7. С. 95-101;URL: https://natural-sciences.ru/ru/article/view?id=37166 (дата обращения: 22.06.2026).
DOI: https://doi.org/10.17513/use.37166