



Потепление климата вызывает продвижение на северо-запад границы распространения многих древесных видов, обуславливает их адаптационные возможности. Липа мелколистная (Tilia cordata Mill.) входит в состав смешанных лесов Евразии, поэтому важно представлять адаптационные особенности этого вида к умеренно холодному климату центра Кольского полуострова. Вопрос о воздействии на широколиственные растения повышенной эмиссии углекислого газа и метана в северном полушарии пока мало изучен и ожидает внимания научного сообщества [1, 2], а генетические свойства видов играют роль в процессе адаптации к изменяющимся климатическим условиям [3].
Влияние климата на форму и асимметрию листовых пластин рассматривалось в различных аспектах [4–8], включая зависимость от ареала древесных видов [9–12]. Установлено, что в северных широтах степень асимметричности увеличивается, а размеры листовых пластин уменьшаются [13–16], например у видов рода Betula отмечена реакция на изменение климатических и других условий в районах высокогорья и на равнине [12].
Как показали предыдущие исследования, популяция липы на Кольском полуострове обладает значительно меньшими линейными размерами листовых пластин и примесью направленной асимметрии (НА) в отличие от подмосковной популяции с высокой флуктуирующей асимметрией (ФА), лишенной НА. Различия в общей асимметричности не выявлены. Форма листовых пластин является контрастирующим параметром и в средней части ареала более разнообразна [17].
Индексы флуктуирующей асимметрии широко используются для определения стабильности популяционного развития растений, включая представителей рода Tilia [18–20]. С использованием линейных признаков изучено влияние поллютантов атмосферы и почвы в урбанизированных районах на увеличение флуктуирующей асимметрии и снижение стабильности развития [21–23]. Однако результаты исследований подвергаются сомнению в связи с частым использованием только одного признака: ширины половины листа, остальные параметры обычно не принимаются во внимание. Ответная реакция у пластических признаков не совпадает по годам, а популяции трудно сравнимы в силу различных условий произрастания.
Пластическая изменчивость (ПИ) величины морфологических признаков часто, но не всегда связана с асимметричностью. В предыдущих исследованиях холодное лето 2017 г. вызвало снижение величины листовых пластин и увеличение флуктуирующей асимметрии, при этом пластическая изменчивость не коррелировала с размером признака [16, 19]. Таким образом, липу мелколистную можно рассматривать как древесную форму факультативных сциофитов с высокой пластической изменчивостью, зависящей от средового воздействия.
В предлагаемой работе изучены морфологические свойства липы мелколистной в популяциях на Кольском полуострове (Мурманская область) и в средней полосе европейской части России (Московская и Владимирская области). Задачей являлось определение изменчивости в сравнительном аспекте четырех географически удаленных популяций. Рабочая гипотеза предполагала статистически значимый эффект влияния климата на асимметричность и форму листовых пластин.
Характеристика мест сбора материала
В Мурманской области в 2021 г. сбор листьев липы мелколистной проведен в двух популяциях. Первая популяция находится на территории Полярно-альпийского ботанического сада-института им. Н.А. Аврорина КНЦ РАН. Это ООПТ с координатами 67°38’ с.ш., 33°58’ в.д. и высотой рельефа 155,5 м над уровнем моря. ПАБСИ располагается в 120 км к северу от Полярного круга вне городской черты г. Апатиты.
Вторая популяция располагается в г. Апатиты, имеет координаты 67°34′03″ с.ш., 33°23′36″ в.д. и высоту рельефа 178 м над уровнем моря. Обе популяции располагаются вне природного ареала распространения липы мелколистной. Описанные провинции характеризуются прохладным и коротким летом со средней температурой 11–14 °С, зимой с температурой -11–-14 °С и годовым количеством осадков 450–560 мм.
В средней полосе России в те же сроки листья липы мелколистной собраны в г. Можайске Московской области с населением 105000 чел., координатами 55°30′ с.ш., 36°02′ в.д. и высотой рельефа 210 м над уровнем моря и в г. Владимире с населением 352000 чел., координатами 56°08′00″ с.ш., 40°25′00″ в.д. и высотой рельефа 150 м над уровнем моря (рис. 1).
В каждом локалитете сбор листовых пластин проведен два раза с перерывом в две недели, по пять листьев с каждого из десяти деревьев со всех сторон нижней части кроны. Всего исследовано 400 листовых пластин примерно одинакового размера. Широтная поясность Кольского полуострова обуславливает более поздние фенодаты появления листьев и зацветания липы мелколистной по сравнению с таковыми средней полосы России.
Материалы и методы исследования
1. Первоначальная обработка и измерение
Листовые пластины после сбора подсушивают на воздухе, помещают в бумажный пресс и сканируют (фотографируют). За экспериментальную единицу принимается как вся популяция, так и листовая пластина. В первом случае находится среднее значение после двукратного сбора, во втором – исходя из совокупности листьев выборки.
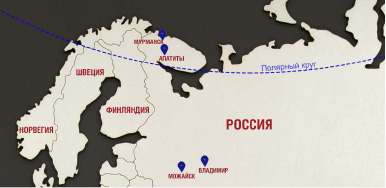
Рис. 1. Места сбора материала
Для измерения использованы 5 линейных признаков: 1 – ширина половины середины листа; 2 – расстояние между основаниями первой жилки первого порядка и второй жилки второго порядка; 3 – расстояние между основаниями второй и третьей жилок второго порядка; 4 – расстояние между основаниями первой и второй жилок первого порядка; 5 – угол между рахисом и первой билатеральной жилкой [19]. Величины линейных признаков (в см) неоднородны по гетерогенности дисперсии, что учитывается при описательной статистике.
В среде Excel проведена первоначальная обработка выборок (R–L), включая определение направленной асимметрии (t-test, H0: L = R) и флуктуирующей асимметрии по формуле FA = |R – L|/(R + L), где R и L – величины правого и левого признаков. Найдены коэффициенты корреляции между ФА различных признаков, а также между величиной (R+L)/2, т.е. величиной признака и величиной ФА для определения аллометрической зависимости «размер листа – ФА». Пластическая изменчивость, как вариация линейных признаков листа, определена по формуле: PL = 1–(x/X), где х и Х – соответственно минимальное и максимальное значения признака. Обработка результатов проведена в среде STATISTICA 10 (StatSoft Ink).
2. Метод геометрической морфометрии
Оценка асимметрии по форме листовых пластин проведена с помощью эталона, усредненного консенсуса, т.е. формы, полученной после усреднения форм всех образцов листьев. Нормализации величин (прокрустовы расстояния) выполнены на основании координат наносимых меток, показывающих отклонение от симметричного консенсуса.
Для нанесения 50 меток по контуру листовых пластин и создания общего файла использован пакет программ TPS (Rholf, 2014), для проведения прокрустового анализа (морфогеометрического дисперсионного анализа) – программа MorphoJ (Klingenberg, 2019). Статистическая значимость оценена на уровне 95 %.
Определена регрессионная зависимость «ФА – величина центроида». Под центроидом понимается величина, равная корню квадратному из суммы квадратов расстояний от меток консенсуса до его геометрического центра. Таким образом, центроид – величина, косвенно служащая для определения размера листовых пластин в геометрическом пространстве. Для определения центров множеств координат меток и расстояний между ними используется ковариационный анализ координат наносимых меток.
Дана оценка влияния расположения популяции, фактора сбора листьев как на форму листовых пластин, так и на их асимметричность. Как показывают предыдущие исследования, ошибка фотографирования крайне мала, поэтому получение изображения проведено один раз, нанесение меток с последующей обработкой проведено дважды. Процедура нанесения настоящих меток первого типа проведена с разбиением левой и правой половин контура на 50 равных отрезков с последующей расстановкой меток [24, 25].
Результаты исследования и их обсуждение
Описательная статистика популяций Кольского полуострова
Условно популяции Кольского полуострова можно обозначить как экспериментальные. Значения линейных признаков в этой крайней северной точке ареала (1,19 см) явно отличаются от таковых в подмосковной и владимирской популяциях (3,87 см) со средним стандартным отклонением соответственно 0,21 и 1,39 (p < 0,05), т.е. размеры листовых пластин здесь примерно в два раза меньше, чем в локалитетах средней полосы России. Тест на направленную асимметрию (t-test, H0:R = L) указывает на ее отсутствие (р > 0,05; табл. 1).
Наибольшее отклонение дисперсии в северных популяциях проявилось по второму признаку, как и в 2020 г., т.е. этот признак наименее пригоден для тестирования ФА мерным способом. Отклонение в эксцессе распределения (R–L) также отмечено по второму признаку (γ = 1,91, ПАБСИ; γ = 8,06; г. Апатиты), по нему же получены и самые высокие значения коэффициента вариации, что, безусловно, влияет на общую величину флуктуирующей асимметрии (0,07 и 0,06 соответственно). Отклонение в распределении (R–L) наблюдается преимущественно в сторону правой половины листа, как и в предыдущий год сбора.
Характерным проявлением выборок ФА является их параметрическое распределение (р > 0,2; K-S test), хотя в предыдущем 2020 г. распределение сильно отличалось от нормального. Четвертый признак внес наибольший вклад в различие. Величина ФА этого признака выше в популяции ПАБСИ (0,078), чем в г. Апатиты (0,051) (табл. 2).
Таблица 1
Свойства распределения (R–L) и величины ФА в популяциях Кольского полуострова
|
Популяция |
Признак |
(R–L) aver |
SD |
s |
(R–L) t-test, t |
λ ±0.24(SD) |
γ ±0.47(SD) |
ФА |
||
|
|R–L|/(R+L) |
K-S, p |
s |
||||||||
|
ПАБСИ |
1 |
0,15 |
2,13 |
4,52 |
0,38 |
0,29 |
0,30 |
0,031±0,002 |
> 0,02 |
0,001 |
|
2 |
-0,04 |
2,77 |
7,69 |
-0,12 |
-0,21 |
1,91 |
0,078±0,007 |
< 0,01 |
0,005 |
|
|
3 |
0,06 |
2,32 |
5,39 |
0,22 |
0,32 |
0,40 |
0,099±0,009 |
< 0,05 |
0,008 |
|
|
4 |
-0,04 |
2,15 |
4,62 |
-0,12 |
-0,24 |
0,03 |
0,078±0,007 |
< 0,05 |
0,005 |
|
|
5 |
0,04 |
1,62 |
2,62 |
0,19 |
-0,09 |
0,03 |
0,061±0,005 |
< 0,01 |
0,002 |
|
|
Апатиты |
1 |
0,13 |
6,32 |
2,51 |
0,37 |
-0,59 |
0,401 |
0,034±0,00 |
< 0,05 |
0,001 |
|
2 |
0,01 |
8,77 |
2,96 |
0,02 |
1,99 |
8,06 |
0,065±0,01 |
< 0,05 |
0,003 |
|
|
3 |
-0,29 |
7,52 |
2,74 |
-1,01 |
-0,74 |
1,99 |
0,092±0,01 |
< 0,05 |
0,006 |
|
|
4 |
-0,12 |
4,30 |
2,07 |
-0,35 |
-0,12 |
2,60 |
0,051±0,01 |
< 0,01 |
0,003 |
|
|
5 |
-0,21 |
2,43 |
1,56 |
-0,98 |
-0,77 |
3,99 |
0,059±0,00 |
< 0,01 |
0,003 |
|
Примечание. (R–L) aver – среднее значение выборки (R–L); SD – стандартное отклонение; λ – асимметричность распределения; γ – эксцесс распределения; test K-S – тест Колмогорова – Смирнова; σ – дисперсия.
Таблица 2
Различие в ФА в двух популяциях Кольского полуострова (двухвыборочный t-test)
|
Признак |
Mean pabsi |
Mean apatity |
t-значение |
df |
p |
|
1 |
0,034 |
0,031 |
0,853 |
207 |
0,394 |
|
2 |
0,065 |
0,078 |
-1,569 |
207 |
0,118 |
|
3 |
0,092 |
0,099 |
-0,643 |
207 |
0,521 |
|
4 |
0,051 |
0,078 |
-3,066 |
207 |
0,002 |
|
5 |
0,059 |
0,061 |
-0,388 |
207 |
0,698 |
|
среднее |
0,060 |
0,069 |
Примечание. mean pabsi – среднее значение (популяция ПАБСИ); mean apatity – среднее значение (популяция г. Апатиты); df – степень свободы; p – уровень вероятности.
Исходя из общего интегрального значения ФА стабильность развития в ПАБСИ (0,07) ниже, чем в г. Апатиты (0,06) (df = 102; t = 1,66; p = 0,02).
Корреляционная зависимость
Корреляция между значениями ФА признаков практически не выявлена. Такая независимость параметров позволяет использовать их величины для определения ФА. Корреляция ФА по величине признака имеет высокие значения лишь по четвертому признаку (r = 0,77) в популяциях средней полосы России. Корреляция ФА–ПИ (r > 0,7) встречается одинаково во всех популяциях (по 12 %).
В 2021 г. в популяции г. Апатиты отмечена связь пластической и флуктуационной изменчивости r = 0,45. На территории ПАБСИ такая связь не выявлена, вероятно, из-за меньшей величины листовых пластин. Очевидно, климатические особенности года влияют на пластическую изменчивость и обуславливают более высокую связь ПИ и ФА. Пластическая изменчивость, особенно в северных популяциях, ассоциирована с флуктуационной изменчивостью.
Геометрическая морфометрия формы листовых пластин
Множество значений координат ХУ в виде меток преобразованы в консенсусную форму для совокупности листовых пластин, удалена составляющая величины, т.е. проведено прокрустово выравнивание. Результаты прокрустова двухфакторного анализа показали изменчивость формы и асимметрии листовых пластин (табл. 3).
Отмечено отсутствие различий в форме листьев (первая строка) как на внутрипопуляционном, так и на межпопуляционном уровнях. Направленная асимметрия (фактор «сторона») различается среди листовых пластин (p < 0,0001). Направленная асимметрия и флуктуирующая асимметрия (взаимодействие обоих факторов) статистически значимы только на уровне листовых пластин при высоком значении степени свободы (df = 14 800).
Ошибка измерения листовых пластин («остатки») составляла 0,50–0,51 % от общей суммы квадратов MS, что считается приемлемым при нахождении флуктуирующей асимметрии. Таким образом, географически популяции различаются по обоим видам асимметрии.
Чтобы определить характер различий, проведен прокрустов анализ для листьев северных популяций и популяций средней полосы России (табл. 4). Направленная асимметрия листовых пластин, как экспериментальной единицы, преобладает на Кольском полуострове («сторона» F = 30,3; p < 0,0001), флуктуирующая асимметрия примерно одинаково проявляется в обеих группах листьев («сторона×лист» F = 3,02 и F = 4,94; p < 0,0001).
На основании координат меток на листовых пластинах для двух регионов проведен ковариационный анализ. Канонические коэффициенты разделились на две группы по форме листовых пластин и по асимметричности (рис. 2).
По первому компоненту формы установлена отрицательная связь с усредненной общей формой листовых пластин и положительная связь асимметрии с дисперсией меток и дисперсией объединенных данных. И асимметрия, и форма отличаются географически по расстоянию в морфо-геометрическом пространстве. Наибольшее прокрустово расстояние между центрами множеств формы установлено для ПАБСИ и г. Можайска (0,09; р < 0,001).
Таблица 3
Географическое различие между северными популяциями и популяциями средней полосы России
|
Фактор |
SS |
MS |
df |
F |
|
Популяция |
0,665 |
0,007 |
100 |
1ns |
|
Сторона |
0,790 |
0,008 |
100 |
1,19 ns |
|
Сбор×сторона |
0,663 |
0,007 |
100 |
64,2*** |
|
Остатки |
14,421 |
0,000 |
139600 |
|
|
Лист |
4,199 |
0,000 |
14800 |
0,96 ns |
|
Сторона |
0,790 |
0,008 |
100 |
26,83*** |
|
Лист×сторона |
4,358 |
0,000 |
14800 |
4,51*** |
|
Остатки |
7,192 |
0,000 |
110200 |
Примечание. SS – сумма квадратов; df – степень свободы; MS – средний квадрат; F – критерий Гудолла; *** – p < 0,0001; ns – статистически не значимо.
Таблица 4
Различия в направленной и флуктуирующей асимметрии
|
Эффект |
Средняя полоса России |
Кольский полуостров |
||||||
|
SS |
MS |
df |
F |
SS |
MS |
df |
F |
|
|
Лист |
1.121 |
0,000 |
8200 |
3,16*** |
3.016 |
0,000 |
6500 |
0,72ns |
|
Сторона |
0.008 |
0,000 |
100 |
1,78*** |
1.946 |
0,019 |
100 |
30,27*** |
|
Лист×сторона |
0.354 |
0,000 |
8200 |
3,02*** |
4.178 |
0,001 |
6500 |
4,94*** |
|
Остатки |
0.712 |
0.000 |
49800 |
7.863 |
0,000 |
60400 |
||
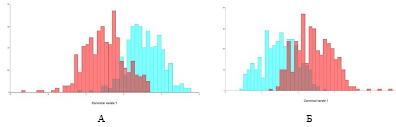
Рис. 2. Распределение частот первого компонента формы (А) и асимметрии (Б). Кирпичный цвет – совокупность северных популяций, лиловый цвет – совокупность популяций средней полосы России
Не все расстояния между множествами асимметрии значимы. Например, г. Владимир и г. Можайск (0,004; р = 0,6), г. Апатиты и ПАБСИ (0,009; р = 0,6) не отличаются по асимметричности. Наибольшее различие установлено между популяциями г. Владимира и ПАБСИ (0,07; р < 0,001), между г. Можайском и ПАБСИ, г. Можайском и г. Апатиты (0,06; р < 0,001). В 2020 г. асимметричность не различалась статистически, так как на нее, по-видимому, влияет год сбора материала.
Как и в сборах предыдущего года, асимметричный компонент формы включает направленную и флуктуирующую асимметрию. Регрессионная связь между величиной консенсуса и ФА не установлена. При работе с линейными признаками замечено проявление прямой связи между ними и ФА. Меньшие размеры листьев в северных популяциях явно связаны с направленной асимметрией. Методом геометрической морфометрии объединяются все метки, проводится разделение на популяционном уровне, регрессионным анализом проверяются аллометрические свойства, т.е. связь между размером консенсуса-центроида и ФА (взаимодействие факторов в прокрустовом анализе).
Направленная асимметрия, как индикатор стресс-фактора, снижающего стабильность развития [22], может проявляться в северных популяциях и служить генетически обусловленной адаптацией к суровым климатическим условиям.
Заключение
Метод геометрической морфометрии с интегральным подходом, позволяющий тестировать изменчивость формы и асимметрии листовых пластин, может быть использован для образцов разного размера. Экспериментальные (северные) популяции обладают значительно меньшими размерами листьев, пластическая изменчивость мерных признаков здесь теснее связана с флуктуационной изменчивостью. Скрытая направленная асимметрия проявляется на биосистемном уровне листовых пластин, но, учитывая стабильность развития как популяционное свойство, правильнее рассматривать асимметричность на популяционном уровне, который в нашем случае указывает на различие в ФА и на снижение стабильности развития в популяциях средней полосы России.
Гипотеза о техногенном влиянии на стабильность развития не подтверждена: на Кольском полуострове в ПАБСИ флуктуирующая асимметрия оказывается выше, чем в популяции г. Апатиты.
Форма листовых пластин является более контрастирующей характеристикой. В средней части ареала форма листовых пластин липы мелколистной более разнообразна.
Библиографическая ссылка
Зыков И.Е., Баранов С.Г., Липпонен И.Н., Полоскова Е.Ю., Кузнецова Д.Д. АДАПТИВНАЯ ИЗМЕНЧИВОСТЬ ЛИСТОВЫХ ПЛАСТИН ЛИПЫ МЕЛКОЛИСТНОЙ (TILIA CORDATA MILL.) // Успехи современного естествознания. 2022. № 7. С. 7-13;URL: https://natural-sciences.ru/ru/article/view?id=37849 (дата обращения: 05.08.2026).
DOI: https://doi.org/10.17513/use.37849