



Введение
Клевер белый (Trifolium repens L.) с его разнообразием фенотипов является отличным индикатором изменений в окружающей среде под воздействием антропогенных факторов. Этот вид растений не только отражает наличие мутаций в своих генах, но и оценивает мутагенную активность почвы, на которой он растет. Исследования в этой области помогают не только контролировать состояние экосистем, но и принимать меры по их восстановлению [1, 2].
Клевер белый для Приморского края является заносным одичавшим растением, которое растет на лугах, лесных опушках, по дорогам, берегам рек, у жилья. Относится к влаголюбивым растениям, хорошо растет при достаточном и даже обильном увлажнении [3].
В течение многих лет выполняется исследование полиморфизма по «седым» пятнам у белого клевера в популяциях г. Уссурийска и Приморского края [4]. Авторами была разработана лабораторная работа «Изучение множественного аллелизма по “седым” пятнам у клевера белого» с использованием исследовательского метода для студентов биологического направления.
Множественный аллелизм представляет собой одно из главных явлений в наследственной изменчивости организма. Разнообразные состояния одного гена возникают в результате мутаций. Поскольку у диплоидных организмов может быть только две аллели из серии множественных аллелей, то в значительной степени увеличивается комбинативная изменчивость организмов. В основе генетического полиморфизма лежит множественный аллелизм, который является неотъемлемым механизмом генетического гомеостаза популяций. В результате популяционный генофонд характеризуется целостностью и сбалансированностью.
Целью исследования стало изучение фенотипического разнообразия в популяциях белого клевера.
Для достижения поставленной цели необходимо решить следующие задачи:
1) ознакомиться с явлением множественного аллелизма, наследованием в серии множественных аллелей;
2) изучить полиморфизм по «седым» пятнам у белого клевера;
3) определить фенотипическое разнообразие популяции белого клевера в городском парке «Зелёный остров» в г. Уссурийске Приморского края;
4) сравнить фенотипическую структуру популяции белого клевера в парке «Зелёный остров» в г. Уссурийске в 2013 г. и в 2023 г.
Материалы и методы исследования
Известно, что полиморфизм – одновременное существование в популяции двух и более резко отличающихся типов организмов. Полиморфизм имеет генетическую основу (генный, хромосомный, внеядерный). Одним из базовых механизмов генетического полиморфизма является множественный аллелизм.
Мутацией определяется переход гена из одного аллельного состояния в другое. Если исходная нормальная аллель А мутирует в рецессивную аллель а, имеет место прямая мутация. В том случае, когда мутантная аллель а превращается в исходную нормальную аллель А, происходит явление обратных мутаций. Однако, как показало исследование генетики аллелей, процесс их мутирования отнюдь не ограничивается взаимопревращением А в а (А → а, а → А). Было выяснено, что мутации могут давать целый ряд разных состояний этого гена (А1, А2, А3, А4...и т.д. и а1, а2, а3, а4...и т.д.) – серию множественных аллелей [5]. Множественный аллелизм – это наличие трех и более аллельных состояний гена. Компаундом называют комбинацию в гетерозиготе двух разных аллелей из набора множественных аллелей. В компаунде одна аллель может доминировать над другой или они могут быть кодоминантны. В ряду аллелей устанавливают и указывают, какая аллель над какой аллелью доминирует [6].
Явление множественного аллелизма встречается в природе и увеличивает полиморфизм популяций, тем самым играет свою роль в процессе эволюции.
У растений по принципу множественного аллелизма функционирует механизм самонесовместимости при опылении. При этом пыльца не может прорастать на рыльцах цветков конкретных генотипов. Гаметофитная самонесовместимость известна у более 10 тысяч покрытосеменных растений и обусловлена генами пыльцевых зерен. Данная система несовместимости определяется у многих растений серией множественных аллелей гена S, (S1, S2, S3, S4, S5, S6 и т.д.) – даже до нескольких сотен у некоторых цветковых растений. Механизм несовместимости работает так: у диплоидных клеток пестика в наличии две аллели из серии; а в гаплоидных пыльцевых зернах в наличии одна аллель гена S. Если в клетках пестика и в пыльцевом зерне содержится одинаковая аллель, то формирование пыльцевой трубки не происходит [7; 8, с. 51].
Множественный аллелизм по гену белоглазия наблюдается и у плодовой мушки. Мушка дрозофила имеет ген белоглазия (white), множественные мутации которого определяют разный цвет глаз в гомозиготе и в гетерозиготе с различной комбинацией мутантных аллелей. Вот некоторые из мутантных аллелей white:
wco – коралловый цвет глаз;
wbl – кровавый;
we – эозиновый;
wch – вишневый;
wa – абрикосовый;
wh – медовый;
wbf – темно-желтый;
wt – слабоокрашенный;
wp – пурпурный;
wi – слоновая кость;
w – белый [8, с. 424].
У многих млекопитающих ген C (color) имеет серии аллелей, например у кроликов:
C – дикий тип, cch – шиншилловая окраска, ch – гималайская окраска, ca – альбинос [8, с. 425].
Множественный аллелизм по группам крови наблюдается у животных и человека. Так, по механизму множественных аллелей наследуются 4 группы крови в системе АВО у людей.
Ген I в системе АВО имеет три различных состояния: IA, IB, Io. Две аллели IA, IB доминируют над третьей аллелью Io, но по отношению друг к другу они равноценны, кодоминантны, обе проявляются фенотипически у гетерозиготы IAIB. Различные генотипы по гену I детерминируют 4 группы крови: I (О) группа – генотип IoIo, II (А) группа – генотипы IAIA или IAIo, III (В) группа – генотипы IBIB или IBIo, IV (АВ) – генотип IAIB [8, с. 427].
У клевера белого, или ползучего (Trifolium repens L.) выявлен генетический полиморфизм по показателю наличия или отсутствия «седого» пятна (называют также светлым, белым пятном) на простых листиках в тройчатом листе, по особой конфигурации пятна. На непохожесть листьев по «седому» пятну влияет серия множественных аллелей гена V, а именно:
V – полное пятно;
VH – полное высокое пятно;
VB – разорванное пятно;
VBH – разорванное высокое пятно;
VP – центральная верхняя точка;
VF – большое сплошное пятно у основания;
VL – низкое треугольное пятно у основания;
v – пятно отсутствует.
Выделяют 11 или больше аллелей данного гена. Кроме 8 общепринятых, авторами учитывается VM – m-образное пятно. В «седом» рисунке можно увидеть доминирование одной из аллелей в компаунде, у некоторых гетерозигот – кодоминирование. Все аллели доминантны по отношению к рецессивной аллели v.
В гистологических исследованиях было установлено, что проявление белого рисунка обусловлено появлением палисадных клеток с небольшим количеством хлоропластов или их отсутствием. Получается, что палисадные клетки уменьшаются в объеме в области рисунка, вытягиваются меньше, межклеточное пространство оказывается больше, чем в зеленой части листа. У других видов клевера есть белые пятна, как у клевера красного, у которого меньше число вариантов пятен. Установлено, что у клевера красного возникновение светлого рисунка зависит от неодинаковой скорости роста палисадных и эпидермальных клеток. В результате палисадные клетки больше подвергаются воздействию солнечного излучения, которое вызывает разрушение хлорофилла [6].
Результаты исследования и их обсуждение
Для изучения полиморфизма рисунка листьев клевера авторы провели сбор материала в популяции на поле в парке «Зелёный остров» в г. Уссурийске в 2023 г. Собирали по одному тройчатому листу с каждого из тысячи растений, далее листья высушивали и наклеивали на гербарную бумагу. Листья клевера пронумеровывали и определяли рисунок на каждом листе. А также использовали гербарные материалы студенческих сборов за 2013 г. Затем провели анализ соотношения разных фенотипов в серии множественных аллелей гена V.
Непосредственно можно видеть только фенотипы, а не генотипы или гены. Генетическую изменчивость определенных популяций чаще всего предпочтительнее выявлять, используя частоты аллелей, а не генотипов, потому что несхожих аллелей чаще всего гораздо меньше, следовательно, генотипов больше. Количество генотипов определяли по формуле:
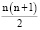 ,
,
где n – число встретившихся в выборке аллелей. Число гомозиготных генотипов соответствует n.
Для расчета числа гетерозиготных генотипов использовали формулу [9, с. 99]:
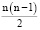 .
.
При статистической обработке данных применяли критерий Стьюдента. Для выведения статистической ошибки использовали общепринятую формулу [9, с. 99].
Количество различных аллелей гена V в популяции в парке в 2023 г. составляет 9, следовательно, в данной популяции
число разных генотипов: 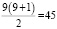 ,
,
число гетерозиготных генотипов: 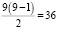 ,
,
гомозиготных генотипов: 9.
Проследим динамику полиморфизма в популяции белого клевера на поле в городском парке «Зелёный остров» в г. Уссурийске от 2013 г. к 2023 г. (таблица).
Частота фенотипов с разными «седыми» пятнами в популяции белого клевера на поле в парке «Зелёный остров» в г. Уссурийске в 2013 г. и 2023 г., %
|
Фенотип |
Популяция 2013 г. |
Популяция 2023 г. |
|
Пятно отсутствует |
9,4±0,9 |
3,7±0,6 |
|
Полное пятно |
36,4±1,5 |
49,6±1,6 |
|
Полное высокое пятно |
9,6±0,9 |
9,7±0,9 |
|
Разорванное пятно |
1,4±0,4 |
3,7±0,6 |
|
Разорванное высокое пятно |
0,3±0,2 |
1,8±0,4 |
|
Центральная верхняя точка |
32,8±1,5 |
16,4±1,2 |
|
Большое сплошное пятно у основания |
0 |
4,1±0,6 |
|
Низкое треугольное пятно у основания |
0,2±0,1 |
0,6±0,2 |
|
m-образное пятно |
9,9±0,9 |
10,4±1,0 |
Примечание: «___» – значения, отличающиеся между выборками 2013 г. и 2023 г. по критерию Стьюдента при p<0,05.
В эти годы наиболее часто встречаются такие фены, как: «полное пятно», «центральная верхняя точка» (таблица).
В 2013 г. отсутствует «большое сплошное пятно у основания».
Наибольшее разнообразие фенов авторы установили в 2023 г. – 9. Меньшее количество фенов обнаружили в 2013 г. – 8 (таблица).
Колебания в частотах установили для таких фенов, как: «пятно отсутствует», «полное пятно», «разорванное пятно», «разорванное высокое пятно», «центральная верхняя точка», «большое сплошное пятно у основания», «низкое треугольное пятно у основания».
Редко встречающимися являются «низкое треугольное пятно у основания» и «большое сплошное пятно у основания». Растения клевера с этими пятнами имеют больше клеток, лишенных хлорофилла, следовательно, такие растения менее жизнеспособны по сравнению с другими.
Итак, полиморфизм по признаку «наличие/отсутствие» «седого» пятна сохраняется в течение 10 лет в популяции на поле в парке «Зелёный остров» в г. Уссурийске.
Выводы современного исследования подтверждаются результатами ранее проводившегося исследования полиморфизма в популяции в парке «Зелёный остров» в 2001 г. в Уссурийске [4].
Ряд исследователей предлагают использовать полиморфизм по белым рисункам на листьях у Trifоlium rеpеns, базирующийся на множественном аллелизме гена V, для биоиндикации загрязнения на территориях произрастания [10, 11]. Предположительно, что на урбанизированных территориях мутагенное действие поллютантов вызывает увеличение количества исследуемых фенов в популяциях Trifоlium rеpеns. Следовательно, фенетический полиморфизм у белого клевера можно использовать для оценки загрязнения окружающей среды [12, 13].
Выводы
В результате проведенной работы авторы пришли к следующим выводам.
1. Показано, что полиморфизм у белого клевера по наличию или отсутствию белых (или так называемых седых) пятен на листьях, по особой конфигурации пятен детерминирован серией из 9 множественных аллелей гена V.
2. Выявлена низкая частота встречаемости фенов «низкое треугольное пятно у основания» и «большое сплошное пятно у основания». Растения с такими пятнами имеют больше клеток, лишенных хлорофилла, следовательно, такие растения менее жизнеспособны.
3. Анализ фенотипической структуры популяции белого клевера на поле в парке «Зелёный остров» в г. Уссурийске показал большее разнообразие фенов в 2023 г. по сравнению с 2013 г. Установлено 9 фенов (соответственно, 45 генотипов) в 2023 г. Установлено 8 фенов (соответственно, 36 генотипов) в 2013 г., отсутствует фен «большое сплошное пятно у основания».
4. Популяция белого клевера в парке «Зелёный остров» в г. Уссурийске характеризуется фенетическим полиморфизмом.
Полученные и обработанные результаты имеют практическое значение в области популяционной фенетики.
Фактический материал может быть использован на лабораторном занятии по генетике «Изучение множественного аллелизма по “седым” пятнам у клевера белого» для студентов биологического направления, а также на уроках биологии в средней школе в разделе «Общая биология» в теме «Генетика». Результаты исследований по изучению полиморфизма белого клевера в природных популяциях используются для проведения экскурсии у студентов и школьников, которая даст возможность продемонстрировать проявление генетически закономерных изменений в природе, находить и констатировать факты, требующие пояснений с позиций генетики, видеть разнообразную изменчивость живых организмов.
Библиографическая ссылка
Шишлова М.А., Быковская Н.В. ИССЛЕДОВАНИЕ ПОЛИМОРФИЗМА У БЕЛОГО КЛЕВЕРА (TRIFOLIUM REPENS L.) В ЗАВИСИМОСТИ ОТ УСЛОВИЙ ПРОИЗРАСТАНИЯ // Успехи современного естествознания. 2024. № 8. С. 43-47;URL: https://natural-sciences.ru/ru/article/view?id=38302 (дата обращения: 08.08.2026).
DOI: https://doi.org/10.17513/use.38302