


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
ECOLOGICAL REGULATION IN POPULATIONS OF SMALL MAMMALS

Динамика популяций мелких млекопитающих формируется под влиянием совокупного действия эндо- и экзогенных факторов и имеет характер сложных сезонных и многолетних колебаний [2–4, 7–9]. Сезонность размножения определяет дискретность популяционной динамики, а поэтому может рассматриваться как один из основных факторов, вызывающих запаздывание реакции популяции на изменения внешних и внутренних условий [4]. Наличие подобного рода запаздывания, высокий репродуктивный потенциал мелких млекопитающих, плотностно-зависимая внутривидовая конкуренция, влияние хищников, изменения кормовой базы и целый ряд других факторов могут являться причинами возникновения, как популяционных автоколебаний, так и хаотических колебаний.
Несмотря на большое количество работ, посвященных динамике популяций и анализу механизмов формирования ее цикличности, в современной экологии не существует единого мнения о причинах и основных факторах возникновения этого явления [1–5, 7–10]. Поддержание численности популяций и механизмы ее формирования – одна из актуальнейших проблем популяционной экологии.
Цель исследования – выявить циклическую составляющую в многолетней динамике популяций рыжей полевки в центре и на периферии ареала и оценить роль внутрипопуляционных и внешних факторов в формировании ее циклов.
Материал и методы исследования
В основу исследования положены материалы многолетнего изучения популяционной динамики на двух стационарных участках, переданные в наше распоряжение А.Д. Бернштейн, А.В. Хворенковым (удмуртский стационар) и Э.В. Ивантером (карельский стационар). Выбор местоположения стационаров осуществлен в соответствии с задачами изучения географических особенностей цикличности рыжей полевки. Удмуртский стационар расположен в бореальной зоне липово-пихтово-еловых подтаежных лесов (57°20′ с. ш., 52° в. д.), он отнесен к оптимальной для рыжей полевки области в центре ареала. Карельский стационар расположен в бореальной зоне, подзоне среднетаежных лесов – периферия ареала рыжей полевки (61°40′ с. ш., 33°30′ в. д.). На территориях обследованных стационаров рыжая полевка доминирует.
Материал собирался стандартным методом ловушко-линий [6]. Отловы проводили регулярно четыре раза в год (апрель, июнь, август и октябрь). За весь период работы отловлено 26893 мелких млекопитающих, в отловах преобладала европейская рыжая полевка (Myodes (Clethrionomys) glareolus Shreber, 1780) – 70,4 % от общего числа отловленных животных (18936). За весь период отработано более 103000 ловушко-суток. Среди большого числа экзогенных факторов анализировались среднемесячные температуры воздуха, количество осадков, высота снежного покрова и зимние корма (семена липы и ели в баллах от 0 до 5). Для обработки данных использовали методы одномерной и многомерной статистики: спектральный и дисперсионный анализы. Различия считались достоверными на уровне значимости р < 0,05.
Результаты исследования и их обсуждение
При рассмотрении многолетней динамики численности полевок возникает вполне закономерный вопрос – случайны колебания ее численности, или они подчиняются некоторым закономерностям. Для решения этого вопроса проведен спектральный анализ многолетних рядов численности рыжей полевки с двух стационарных географически удаленных участков.
В спектрах рядов численности, обеих популяций выявлен явно выраженный пик спектральной плотности соответствующий периоду от 2 до 5 лет. На спектрограмме обнаружен еще один мощный выброс соответствующий сезонному пику с периодом один год. Поэтому можно считать, что наблюдаемые изменения численности популяций рыжей полевки представляют собой сложный процесс, состоящий из суммы двух колебаний (сезонной и многолетней составляющих). В исследованных популяциях выделены четыре фазы популяционного цикла. Кроме того, обнаружены повторяющиеся переходы из одной фазы в другие, но для каждой популяции характерна присущая только ей структура переходов. На удмуртском стационаре (центр ареала) популяционный цикл имеет продолжительность 2–4 года, а на карельском стационаре (периферия ареала) длительность популяционного цикла колеблется от 2 до 5 лет.
В удмуртской и карельской популяции, несмотря на постоянно меняющиеся условия каждого конкретного года, во всех случаях происходит переход из фазы «пик» в фазу «депрессия». Динамика численности и структуры населения наблюдаемых популяций более всего напоминает один из типичных сценариев перехода хаотического движения через некоторые промежуточные к стационарному состоянию. При таком характере динамики всплески хаотического поведения чередуются с участками, на которых происходят почти периодические колебания.
В результате проведенного анализа сценарий развития циклов рыжей полевки можно представить себе следующим образом. Популяционный цикл представляет собой на две качественно различные части. Первая – «популяционный крах» или «детерминированная» часть цикла – переход из фазы «пик» в фазу «депрессия». Этот переход наблюдается в обеих популяциях в разные годы, несмотря на все многообразие условий каждого конкретного из них и местонахождения популяции. Такая ситуация может иметь место только в том случае, если демографические процессы определяются главным образом внутрипопуляционными факторами и слабо зависят от внешних условий.
Вторую часть цикла можно назвать «стохастической», она начинается с устойчивой точки цикла фазы «депрессия». Длительность этой части цикла связана с возможностью популяции увеличивать свою численность, а скорость этого процесса определяется различным соотношением влияния как внешних, так и внутрипопуляционных факторов. Вероятнее всего, определяющими в этой части цикла являются внешние условия (погодные и кормовые факторы, численность хищников, месторасположение популяции в ареале вида и др.). Подобное явления отмечено и для других частей ареала рыжей полевки.
Каким образом можно объяснить феномен цикличности или в общем случае процесс регулирования численности? На рисунке представлена общая схема регулирования численности в системе кормовой ресурс – мелкие млекопитающие – хищники. В данном случае к внешним условиям откосится весь комплекс погодных и климатических факторов (температура, количество осадков, влажность, антропогенные факторы), кормовой ресурс и хищники, а к внутренним внутрипопуляционные гомеостатические плотностно-зависимые механизмы регуляции.
Согласно современным представлениям схему регулирования численности мелких млекопитающих в общем виде можно представить состоящей из следующих основных блоков (рисунок): грызуны, хищники, кормовые ресурсы (растительный покров) и комплекс внешних условий [2–5, 7, 8].
Численность популяций полевок определяет ее плотность и структуру населения, которые в зависимости от количества и качества кормовых ресурсов и численности хищников могут «включать» или «выключать» плотностно-зависимые механизмы изменяющие уровень смертности и интенсивность размножения зверьков. Увеличение плотности зверьков на территории приводит к уменьшению густоты растительного покрова и снижению наземной и корневой биомассы растений и если эти воздействия превышают величины допустимого воздействия на растительные сообщества, уменьшается его продуктивность, что в дальнейшем может стать причиной падения численности.
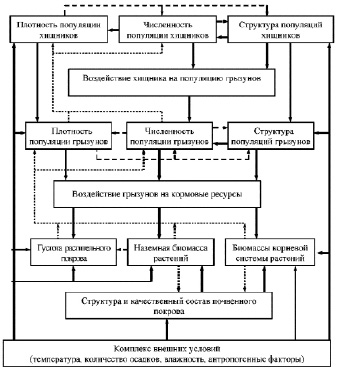
Схема взаимодействия популяций грызунов, хищников и кормовой базы. Примечания: сплошная линия – прямое воздействие; линия из точек – обратные связи; пунктирная линия – взаимодействия на одном трофическом уровне
Известно, что переход биомассы с уровня первичных продуцентов на уровень первичных консументов связан с потерей вещества и энергии. В среднем считается, что лишь 10 % фитомассы и связанной с ней энергии переходит с предыдущего уровня на следующий, остальная энергия и биомасса расходуется на подержание жизнедеятельности и процессы роста и развития. В лесных биоценозах кормовые ресурсы в летний период настолько велики, что изъятие 10 % общей биомассы не могут стать причинной снижения численности полевок. В то время как высокая урожайность семена липы и ели осенью предыдущего года (зимние корма) могут инициировать зимнее размножение и высокую весеннюю численность следующего года [4].
Численность хищников также как и численность полевок формирует свою плотность и структуру популяции. Основой пищевого рациона хищников служат мелкие млекопитающие. Поэтому увеличение плотности и изменение структуры населения хищников приводит к возрастанию потребностей животных в пище, что может приводит к снижению плотность популяций грызунов. Но согласно опубликованным ранее данным хищники не могут являться причиной снижения численности полевок, так как они могут изъять из популяции полевок не более 5–7 % от общей биомассы полевок, не оказывая тем самым существенного влияния на популяцию своих жертв [2].
Заключение
Таким образом, внутрипопуляционные факторы формируют «детерминируемую» часть цикла. Переход этот для обеих популяций осуществляется за один осенне-зимне-весенний период и не влияет на длительность цикла. Длительность «стохастической» части цикла определяется с одной стороны состоянием популяции и ее возможностью увеличить свою численность, а с другой ограничивающими факторами – погодные и кормовые условия, пресс хищников и др., в число которых входит и месторасположение популяции в ареале вида. В центре ареала длительность «стохастической» части цикла составляет 1–3 года. На периферии ареала этот промежуток времени составляет 2–4 года.
Автор выражает благодарность Трушину С.П. за помощь в подготовке статьи.
Работа выполнена при финансовой поддержке программы научных исследований УрО РАН (проект № 12-С-4-1012) и РФФИ (грант № 13-04-00802).
Библиографическая ссылка
Жигальский О.А. ЭКОЛОГИЧЕСКОЕ РЕГУЛИРОВАНИЕ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ МЕЛКИХ МЛЕКОПИТАЮЩИХ // Успехи современного естествознания. 2013. № 2. С. 61-64;URL: https://natural-sciences.ru/en/article/view?id=31365 (дата обращения: 07.07.2026).