


Scientific journal
Advances in current natural sciences
ISSN 1681-7494
"Перечень" ВАК
ИФ РИНЦ = 0,976
ENTROPIC APPROACH FOR THE EVALUATION OF THE ORDER OF CHAOS AND BIOLOGICAL SYSTEMS

Концепция структурных уровней живых организмов позволяет расположить их в иерархическом соподчинении. Согласно критерию масштабности, биологический уровень включает подуровни: макромолекул (нуклеиновые кислоты, ДНК, РНК, белки), клеточный уровень, микроорганический (одноклеточные организмы), органов и тканей организмов в целом, популяционный, биоценозный и биосферный.
Живые организмы образуются из всевозможных малых органических молекул – мономеров, которые при объединении создают макромолекулы. Особую роль играют три класса мономеров: аминокислоты, нуклеотиды и моносахариды. Это те «кирпичики», из которых затем строятся полимерные макромолекулы – белки, нуклеиновые кислоты и полисахариды. Белки – это органические соединения, входящие в состав всех живых организмов, состоящие из большого числа мономеров. В состав белков входят 20 стандартных аминокислот. Свойства белков определяются пространственной трехмерной структурой их цепей. Изменяя в цепи лишь одну аминокислоту, можно получить молекулу с совершенно другой структурой и иными свойствами. Огромное разнообразие живых организмов на нашей планете определяется различиями в составе и пространственной форме составляющих их белков. Нуклеотиды – это мономерные звенья цепи нуклеиновых кислот. Они представляют собой химические соединения остатков трех веществ: азотистого основания, углевода и фосфорной кислоты. Существует два вида нуклеиновых кислот – дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК). ДНК содержит генетическую информацию о последовательности аминокислот в полипептидных цепях и определяет структуру белков. РНК отвечает за создание белков. Оказалось, что молекула ДНК состоит из двух мономерных цепей, идущих в противоположных направлениях и спирально закрученных одна вокруг другой. ДНК всего органического мира образованы соединением четырех видов нуклеотидов- аденин (A), гуанин (Г), цитозин (С) и тимин (Т), их расположение в молекуле ДНК дает указание молекуле РНК, как надо строить белок. Молекула РНК одноцепочная. Структура РНК также создается чередованием четырех типов нуклеотидов, но их состав несколько отличается от состава таковых в молекулах ДНК тем, что вместо тимина (Т) используется урацил (U) [1].
В биологических системах могут идти пpоцессы как с возpастанием, так и уменьшением энтpопии. Понятие энтропии имеет множество трактовок в самых разнообразных областях человеческих знаний. Наряду с энтропией Клаузиуса появилась статическая, информационная, математическая, лингвистическая, интеллектуальная и другие энтропии. Энтропия стала базисным понятием теории информации и стала выступать как мера неопределенности некоторой ситуации. Для характеристики меры сложности системы У. Эшби [11] впервые предложил использовать понятие энтропии. Система взаимодействует с внешним миром как единое целое. В целом же, система не теряет своей организованности или высокой упорядоченности. Открытые системы могут обмениваться с окружающими телами энергией, веществом и, что не менее важно, информацией. Чтобы биологическая система действовала и взаимодействовала со средой, она должна потреблять информацию из среды и сообщать информацию среде. Этот процесс называется информационным метаболизмом, который совместно с вещественным и материальным метаболизмом образует полный метаболизм. Впервые понятия энтропия и информация связал Шеннон [7]. С его подачи энтропия – это количество информации, приходящейся на одно элементарное сообщение источника, вырабатывающего статистически независимые сообщения. Получение какого-либо количества информации равно потерянной энтропии. Информационная энтропия для независимых случайных событий x с N возможными состояниями рассчитывается по формуле
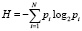 ,
,
где Pi – вероятность частоты встречаемости некоторого события.
Впервые для оценки степени структурированности биоценозов Мак-Артур в 1955 г. использовал общее уравнение энтропии Шеннона [12], в котором, pi заменил на pi = ni /N; (где ni – общее число особей вида i, N – общее число особей во всем биоценозе). В 1957 г. Р. Маргалеф постулировал теоретическую концепцию, согласно которой разнообразие соответствует энтропии при случайном выборе видов из сообщества [13]. В результате этих работ большое распространение и повсеместное признание получил индекс Шеннона Н, иногда называемый информационным индексом разнообразия К. Шеннона [3].
При расчете энтропии Н по Шеннону считается, что каждая проба – случайная выборка из сообщества, а соотношение видов в пробе отражает их реальное соотношение в природе. В качестве оценок вероятностей независимых событий рi для формулы (1) могут быть использованы следующие апостериорные отношения: удельная численность i-го вида, как частное от деления его численности ni на общую численность всех видов N, взятых для анализа (pi = ni /N):
H = –Σni /N log2 (ni/N).
Использование индекса разнообразия Шеннона в экологии и, в частности, для оценки степени структурированности популяций и биоценозов очень детально обсуждается в работах [3, 8, 9]. Особенности функционирования биологических систем разного уровня с точки зрения теории информации были рассмотрены также в работе [10]. Шмальгаузеном были введены понятия о каналах прямой и обратной связи, по которым передается генетическая и фенотипическая информация, рассмотрены закономерности преобразования и кодирования биологической информации.
Целью данной работы является применение индекса Шеннона к биологическим системам, включающим подуровни: макромолекул (нуклеиновые кислоты, белки и РНК) и клеточный уровень.
В последние годы для комплексной оценки качества поверхностных вод сотрудниками кафедры экологической химии ЕГУ используется энтропийный индекс качества воды – G, который получается из индекса Шеннона [4]. G-функция характеризует гидроэкологические системы со стороны соотношения порядка и хаоса, мерами которых являются геоэкологическая синтропия – I [5] и энтропия Шеннона H, соответственно G = H/I. Значения G-функции говорят о том, что и в какой мере преобладает в структуре системы: хаос или порядок. Так, если G ˂ 1, то в структуре системы преобладает порядок, в противном случае, когда G ˃ 1, – хаос. При G = 1 хаос и порядок уравновешивают друг друга, и структурная организация системы является равновесной.
Методика расчета. Для расчета значений I, H и G пользуемся следующим вычислительным алгоритмом:
1. Определяется число элементов в системе – n.
2. Далее оценивается общая сумма элементов: N = ∑n.
3. Потом вычисляются: log2N, nlog2n и ∑ nlog2n.
4. Рассчитывается геоэкологическая синтропия [11] (I) и энтропия (H):
I = ∑ nlog2n / N и H = log2N – I.
5. После чего определяется индекс качества воды: G = H/I.
В соответствии с целью работы и постановки задачи произведены расчеты функции белковых молекул, РНК, крахмала и живого тела.
Первичные структуры, белковые молекулы и РНК представляют собой, соответственно, последовательности аминокислотных остатков и нуклеотидов, и могут рассматриваться как системы, разделенные на части по одинаковым видам соответствующих аминокислот и нуклеотидов. Например, первичная структура молекулы глюкагона следующая:
His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu-Met-Asn-Thr
Для глюкагона N = 29, число нуклеиновых кислот равно 15, а число одинаковых остатков (n) изменяется от 1 до 4. Соответствующие расчеты функций I, H и G приведены в табл. 1, откуда следует, что цепь глюкагона относится к хаотичному типу систем.
В табл. 2 даны значения I, H и G для белковых молекул.
Из табл. 1 и 2 видно, что аминокислотные цепи глукагона и кортикотропина обладают явной хаотичной структурой (G ˃ 1). Цепи же всех остальных белковых молекул относятся к синергетичному типу систем и проявляют тенденцию к уменьшению и приближению G = 1 по мере роста в белковой цепи общего числа аминокислотных остатков.
Приведем некоторые статистические данные по геному вируса HIVH3CG [6]. Расчеты приведены в табл. 3.
Так как для РНК G ˂ 0,2, то в структуре системы преобладает порядок.
В клетках живых систем найдены многие химические элементы, присутствующие в окружающей среде, однако необходимы для жизни лишь около 20 из них. Эти элементы получили название биогенных.
Таблица 1
Расчет значений I, H и G в структуре молекули глюкагона
|
№ п/п |
Аминокислота |
n |
nlog2n |
№ п/п |
Аминокислота |
n |
nlog2n |
|
1 |
Gly |
1 |
0 |
9 |
Leu |
2 |
2 |
|
2 |
Ala |
1 |
0 |
10 |
Phe |
2 |
2 |
|
3 |
Val |
1 |
0 |
11 |
Tyr |
3 |
4,76 |
|
4 |
Lys |
1 |
0 |
12 |
Asp |
3 |
4,76 |
|
5 |
His |
1 |
0 |
13 |
Gln- |
3 |
4,76 |
|
6 |
Met |
1 |
0 |
14 |
Thr |
3 |
4,76 |
|
7 |
Asn |
1 |
0 |
15 |
Ser |
4 |
8 |
|
8 |
Arg |
2 |
2 |
||||
|
N = 29 ∑ nlog2n = 33,1 I = 26:21 = 1,14 H = log229 – 1,14 = 3,75 G = 3,75:1,14 = 3,29 |
|||||||
Таблица 2
Значения N, I, H и G белковых молекул
|
Белковые молекулы |
N [12] |
I |
H |
G |
|
Глюкагон |
29 |
1,14 |
3,75 |
3,29 |
|
Кортикотропин |
39 |
1,45 |
3,84 |
2,65 |
|
Цитохром с |
104 |
2,75 |
3,95 |
1,74 |
|
Миоглобин |
152 |
3,30 |
3,95 |
1,20 |
|
Лютеинизирующии гормон |
205 |
3,57 |
4,11 |
1,15 |
|
Тромбин |
259 |
3,81 |
4,21 |
1,10 |
|
Фосфорилаза гликогена |
841 |
5,54 |
4,18 |
0,75 |
Таблица 3
Расчет I, H и G генома вируса (HIVH3CG)
|
Нуклеотиды |
n |
nlog2n |
|
U |
2168 |
24026,1 |
|
C |
1785 |
19281,1 |
|
A |
3432 |
40308,3 |
|
G |
2364 |
26493,4 |
|
I = Σ nlog2n:N = 110108,9:9749 = 11,29 H = log2N – I = log29749 – 11,29 = 1,95 G = H/I = 1,95:11,29 = 0,173 |
||
Таблица 4
Расчет I, H и G клетки
|
Элементы |
n |
nlog2n |
|
C |
18 |
75,0 |
|
H |
10 |
33,2 |
|
O |
70 |
428,8 |
|
N |
2 |
2 |
|
I = Σ nlog2n:N = 539:100 = 5,39 H = log2N – I = log2100 – 5,39 = 1,25 G = H/I = 1,25:5,39 = 0,232 |
||
В среднем около 70 % массы организмов составляет кислород, 18 % – углерод, 10 % – водород, и 2 % – остальные элементы (азот, фосфор, калий, кальций, сера и т.д.) [10].
Из табл. 4 следует, что в структуре клетки также преобладает порядок.
Таким образом, впервые с помощью энтропийного индекса Шеннона проведен структурный анализ состояния биологических систем на уровне белков, рибонуклеиновой кислоты, крахмала и клетки. Установлено, что в ряду: белок → РНК → клетка энтропия уменьшается, а геоэкологическая синтропия растет. Для генома вируса HIVH3CG и клетки G ˂ 0,2, что свидетельствует о высокой степени свободы и порядка этих систем. Для белка G-функция стремится к единице, что свидетельствует о том, что структурная организация системы является равновесной.
Библиографическая ссылка
Симонян Г.С., Симонян А.Г. ЭНТРОПИЙНЫЙ ПОДХОД К ОЦЕНКЕ ХАОСА И ПОРЯДКА БИОЛОГИЧЕСКИХ СИСТЕМ // Успехи современного естествознания. 2015. № 9-1. С. 130-133;URL: https://natural-sciences.ru/en/article/view?id=35544 (дата обращения: 16.07.2026).