



Условием успешного, устойчивого существования лесных экосистем является определенная сбалансированность между показателями продукционных и деструкционных процессов. При этом главный вклад в деструкционные процессы вносят грибы, в частности, ксилотрофные [1. 6]. Функции, выполняемые дереворазрушающими грибами, оказывают прямое влияние на лесные экосистемы, участвуя в круговороте веществ посредством разложения лигнин-целлюлозных соединений и продуцирования за счет этого собственной биомассы. Эти же функции грибов оказывают косвенное влияние на жизнедеятельность человека, обеспечивая существование лесных экосистем, являющихся источником кислорода, а также источником многочисленных древесных и недревесных продуктов, используемых человеком.
Большое значение для анализа деструкционных процессов, происходящих в экосистемах, имеет не только видовой состав редуцентов, но и их количественные показатели. Из общего числа видов любого трофического уровня или сообщества в целом обычно лишь некоторые доминируют, т.е. имеют относительно большую значимость (большую численность, продуктивность). Количественная сторона представленности различных видов ксилотрофных грибов в микоценозах является индикатором их экосистемной роли, т.е. позволяет выявить те виды, которые играют ведущую роль в процессе микогенной деструкции древесных остатков в данном биоценозе, являются доминирующими видами, слагающими ядро той или иной формационной микобиоты.
С целью анализа функциональной структуры микоценозов дереворазрушающих грибов, нами были проанализированы данные учета количества плодовых тел грибов этой трофической группы, собранные в результате многолетних исследований, проведенных в 1994-2012 гг. в Южном Приуралье в пределах административных границ Оренбургской области. В ходе исследований были изучены сообщества грибов лесов разного генезиса в лесостепной и степной зонах региона. Исследования проводились по классической методике учета плодовых тел [1].
Выявление доминирующих видов в микоценозах ксилотрофных грибов затруднено по ряду причин: в первую очередь – из-за трудности учета их действительной численности в биоценозе, так как количественный учет производится по базидиомам, многие из которых подвержены быстрому разрушению насекомыми и другими экзогенными факторами и, кроме того, их численность не всегда соответствует роли данного вида в процессе разложения древесины в том или ином биоценозе [4]. В.А. Мухин [1] предложил использовать в качестве показателя значимости того или иного вида гриба в биологическом разложении древесины положение данного вида в иерархической структуре численности видов, связанных с древесиной определенного вида древесного растения. Проведенный нами анализ численности видов дереворазрушающих грибов в микоценозах, как мы и предполагали, показал, что в большинстве случаев максимальную численность имеют один-два вида, а прочие характеризуются относительно низкой встречаемостью(рис. 1).
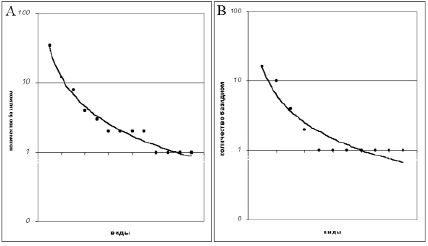
Рис. 1. Относительная значимость видов в микоценозах мелколиственных лесов Бузулукскогорайона. Микоценозы: А – березняка, В – осинника
Соответственно невелико и количество доминирующих видов грибов деструкторов древесины в микоценозах Южного Приуралья. К ним относятся Trichaptum fuscoviolaceum (Ehrenb.:Fr.) Ryv., Deadalea quercina (L.:Fr.) Pers., Fomitoporia robusta (P.Karst.) Fiasson & Niemela, Inocutis dryophila (Berk.) Fiasson & Niemela, Fomes fomentarius (L.: Fr.) Fr.. Также можно выделить ряд видов – кодоминантов (табл. 1).
Таблица 1
Участие доминантов и кодоминантов в сообществах ксилотрофных грибов
|
ВИДЫ |
Доля древесных остатков, на которых отмечен вид ( %)* |
||
|
I |
II |
III |
|
|
1 |
2 |
3 |
4 |
|
Микоценозы дубняков |
|||
|
Daedalea quercina |
42,23 |
28,77 |
34,54 |
|
Hymenochaete tabacina |
21,75 |
12,50 |
18,20 |
|
Fomitoporia robusta |
18,71 |
25,42 |
18,2 |
|
Inocutis dryophila |
17,31 |
24,61 |
14,45 |
|
Микоценозы вязовников |
|||
|
Fomes fomentarius |
12,41 |
14,83 |
21,43 |
|
Polyporus squamosus |
16,92 |
10,08 |
43,33 |
|
Auricularia mesenterica |
26,15 |
2,03 |
28,75 |
|
Микоценозы кленовников |
|||
|
Fomes fomentarius |
25,54 |
10,84 |
16,00 |
|
Polyporus squamosus |
6,41 |
2,48 |
28,18 |
|
Schizophyllum commune |
13,50 |
11,92 |
10,09 |
|
Микоценозы липняков |
|||
|
Schizophyllum commune |
39,32 |
28,92 |
24,54 |
|
Fomitopsis pinicola |
13,64 |
18,13 |
– |
|
Stereum hirsutum |
8,23 |
23,77 |
23,80 |
|
Микоценозы ольшаников |
|||
|
Inonotus radiatus |
24,90 |
22,04 |
28,57 |
|
Phellinus alni |
9,12 |
19,64 |
9,52 |
|
Fomitopsis pinicola |
16,87 |
18,75 |
- |
|
Микоценозы березняков |
|||
|
Fomes fomentarius |
42,52 |
40,18 |
33,96 |
|
Piptoporus betulinus |
12,27 |
10,26 |
9,79 |
|
Stereum subtomentosum |
13,65 |
9,10 |
22,49 |
|
Микоценозы осинников |
|||
|
Fomes fomentarius |
34,25 |
28,22 |
35,52 |
|
Phellinus tremulae |
22,89 |
29,14 |
19,64 |
|
Pleurotus calyptratus |
13,24 |
16,67 |
31,45 |
|
Inocutis rheades |
13,33 |
15,91 |
15,05 |
* – I – лесостепная зона; II – леса южных отрогов Уральских гор; III – леса степной зоны.
Полученные данные позволяют утверждать, что комплекс доминантных видов в отдельных микоценозах формационных биот достаточно постоянен, однако относительная значимость каждого из этих видов зачастую заметно варьирует.
Исходя из вышесказанного, в биоте ксилотрофных базидиомицетов Южного Приуралья можно выделить четыре комплекса доминирующих видов. Первый из них характерен для сосняков и сформирован Trichaptum fuscoviolaceum и рядом других видов, таких как Lentinus lepideus (Fr.: Fr.) Fr. и видов рода Postia.
Второй комплекс формируется стенотрофными «кверцетальными» видами, такими как Daedalea quercina, Inocutis dryophila, Fomitoporia robusta, Hymenochaete tabacina. В каждом из изученных микоценозов дубняков присутствовал хотя бы один из этих видов и в большинстве случаев он являлся доминантом. При этом от микоценоза к микоценозу наблюдается изменение роли отдельных видов – от доминанта до кодоминанта второго порядка (см. табл. 1).
Третий комплекс характерен для ольшаников и сформирован Inocutis radiatus (Sow.: Fr.) P. Karst. с участием ряда других видов. Этот вид является доминантом более 60 % изученных микоценозов ольшаников. В лесах лесостепной зоны к нему присоединяются кодоминанты Phellinus alni (Bond.) Parmasto, Stereum subtomentosum Pouzar; в предгорьях Южного Урала – кодоминанты Fomes fomentarius и Fomitopsis pinicola.
Четвертый комплекс образован Fomes fomentarius при участии ряда других видов и характерен для микоценозов мелколиственных и кленово-вязовых лесов (рис. 2).
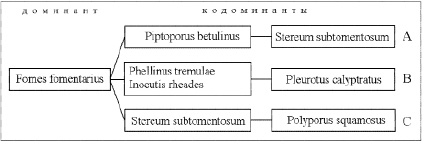
Рис. 2. Структура комплекса доминирующих видов, образованного Fomes fomentarius. Микоценозы: А – березняков, В – осинников; С – кленово-вязовых лесов
Спектр доминантных видов мелколиственных лесов достаточно однотипен и включает, помимо Fomes fomentarius, кодоминант Piptoporus betulinus (Bull.:Fr.) P.Karst.. В пределах степной зоны в микоценозах березняков несколько выше численность Stereum subtomentosum; в микоценозах осинников возрастает значение Pleurotus calyptratus (Lindbl.) Sacc.
Для микоценозов кленово-вязовых лесов характерна полидоминантность с некоторым преобладанием Fomes fomentarius. К числу кодоминантов могут быть отнесены Stereum subtomentosum, Schizophyllum commune. В низкогорных кленово-вязовых лесах в роли кодоминанта выступает Polyporus squamosus. В отдельных микоценозах в комплекс доминирующих видов также входят Cerrena unicolor, Fomitopsis pinicola, Crepidotus mollis.
Микоценозы липняков несколько отличаются от сообществ грибов кленовников и вязовников: в них доминантом является Schizophyllum commune, кодоминантами – Fomitopsis pinicola и Panellus stipticus.
Сравнение комплексов доминантных видов ксилотрофных базидиомицетов в Южного Приуралья и Западно-Сибирской равнины [1] показывает, что виды Fomes fomentarius, Phellinus tremulae, Piptoporus betulinus, Trichaptum fuscoviolaceum, являющиеся доминантами в Западной Сибири, сохраняют свои позиции и в сообществах дереворазрушающих грибов Южного Приуралья. Таким образом, ведущую роль в процессах деструкции древесины в лесах указанных регионов играют одни и те же широко распространенные виды. Это можно расценивать, как свидетельство их значительной экологической пластичности.
Существует и ряд принципиальных отличий между комплексами доминантных видов микоценозов этих территорий. Так, в березовых лесах Западной Сибири Fomes fomentarius является доминантным видом в широком диапазоне – от северной тайги до подтайги, однако в лесостепи уступает свое место Daedaleopsis confragosa. Этот вид часто встречается в микоценозах березняков Южного Приуралья, однако его относительная значимость в микоценозах невелика (3-15 %). Причинами некоторого варьирования состава видов, играющих ведущую роль в процессах деструкции древесины в лесах Западно-Сибирской равнины и Южного Приуралья, являются географическое распространение активных видов, определяющееся их экологической пластичностью, а также упоминавшаяся выше сложность полевого определения объективной роли того или иного вида в микоценозе.
Значимость видов в сообществе может быть выражена с помощью индекса доминирования Симпсона [3]. Для микоценозов ксилотрофных базидиомицетов Южного Приуралья характерны относительно невысокие показатели концентрации доминирования (средняя величина – 0,21; медиана – 0,19; варьирование от 0,04 до 0,63). В этом плане изученные микоценозы несколько опережают сообщества ксилотрофных грибов Западно-Сибирской равнины, для которых максимальное значение индекса Симпсона составляет 0,40 [1].
В широтном градиенте наблюдается тенденция к некоторому увеличению концентрации доминирования в микоценозах (табл. 2). Минимальные показатели характерны для сообществ наиболее увлажненных местообитаний – пойменных лесов и лесов предгорий Южного Урала; максимальные показатели характерны для микоценозов степных лесов. При этом дисперсии в пределах всех указанных районов невелики.
Таблица 2
Варьирование концентрации доминирования по стациям
|
Стации |
m ± M |
Lim |
σ2 |
|
Низкогорные леса |
0,18 ± 0,01 |
0,04 – 0,43 |
0,006 |
|
Леса лесостепной зоны |
0,23 ± 0,02 |
0,06 – 0,63 |
0,017 |
|
Леса степной зоны |
0,24 ± 0,02 |
0,08 – 0,43 |
0,013 |
|
Пойменные леса |
0,18 ± 0,03 |
0,06 – 0,32 |
0,007 |
|
Общее: |
0,21 ± 0,01 |
0,04 – 0,63 |
0,011 |
Отмеченная тенденция вполне согласуется с принятым в экологии подходом к роли доминирующих видов в сообществах, согласно которому доминирование видов лучше выражено в сообществах, обитающих в средовых экстремумах [3]. Уровень доминирования этих видов в широтном градиенте в большинстве случаев возрастает. Это связано с общим сокращением видового разнообразия сообществ дереворазрушающих грибов в степной зоне за счет элиминации видов с малой экологической пластичностью, что приводит в конечном итоге к снижению тактической устойчивости сообществ [2].
Отличия в степени доминирования наблюдаются и между формационными микобиотами. Максимальное доминирование характерно для сообществ, относящихся к формационным микобиотам тополевников, дубняков, березняков (среднее значение – 0,26, 0,24 и 0,23 соответственно). При этом в пределах каждой из формационных микобиот концентрация доминирования испытывает локальные изменения (табл. 3).
Таблица 3
Средние показатели концентрации доминирования в формационных микобиотах
|
Стации |
Формационные микобиоты |
|||
|
березняки |
осинники |
дубняки |
кленовники и вязовники |
|
|
Низкогорные леса |
0,22 |
0,21 |
0,18 |
0,10 |
|
Леса лесостепной зоны |
0,24 |
0,22 |
0,31 |
0,14 |
|
Леса степной зоны |
0,23 |
0,23 |
0,16 |
0,24 |
|
Пойменные леса |
– |
0,12 |
0,13-0,32 |
0,12 |
|
Среднее |
0,23 |
0,21 |
0,24 |
0,14 |
Анализ изменения степени доминирования в формационных микобиотах по районам показал, что ее увеличение в широтном градиенте хорошо прослеживается только для микобиоты кленово-вязовых лесов. В микобиотах мелколиственных лесов данный показатель варьирует очень слабо. Это может быть связано с тем, что эти леса в пределах степной зоны преимущественно приурочены к максимально увлажненным местам, что несколько сглаживает экстремальность условий этих местообитаний для грибов. Для микобиоты дубрав характерно снижение концентрации доминирования от дубрав лесостепи к остепненным дубнякам Общего Срыта. Можно предположить, что значительная выраженность доминирования в этих лесах связана с высоким уровнем антропогенной нагрузки (рубки), который обусловливает доминирование в микоценозах как фитопатогенных (Inocutis dryophila, Fomitoporia robusta), так и специфических сапротрофных видов (Daedalea quercina).
Минимальная величина индекса Симпсона (0,04-0,10) была отмечена в микоценозах низкогорных вязовников и кленовников.
Таким образом, в биоте ксилотрофных базидиомицетов Южного Приуралья присутствует ограниченное число видов, играющих ведущую роль в процессе биодеструкции. Эти виды являются одними из наиболее значимых индикаторов функционирования микобиоты и, следовательно, важным элементом ее ресурсного потенциала [5]. К ним относятся как специфичные виды, являющиеся эдификаторами определенных лесных формаций (Daedalea quercina, Inocutis dryophila, Inocutis rheades, Inonotus radiatus, Fomitoporia punctata, Phellinus tremulae, Piptoporus betulinus), так и эвритрофные виды, характеризующиеся широкой экологической амплитудой (Fomes fomentarius, Schizophyllum commune, Stereum subtomentosum и др.). Значение этих видов очень велико, поскольку они обеспечивают разложение древесных остатков во многих лесах области (например Fomes fomentarius является доминантом или кодоминантом в 42,3 % изученных микоценозов).
Исследование функциональной структуры сообществ на основе учета количественных показателей может стать эффективным инструментом оценки состояния микобиоты и лесных экосистем в целом.
Библиографическая ссылка
Сафонов М.А., Каменева И.Н., Булгаков Е.А. ФУНКЦИОНАЛЬНАЯ СТРУКТУРА СООБЩЕСТВ ДЕРЕВОРАЗРУШАЮЩИХ БАЗИДИАЛЬНЫХ ГРИБОВ ЮЖНОГО ПРИУРАЛЬЯ // Успехи современного естествознания. 2013. № 10. С. 222-227;URL: https://natural-sciences.ru/ru/article/view?id=33108 (дата обращения: 17.07.2026).